Molecular Biology 5thRobert F WeaverChap 13
Bạn đang xem bản rút gọn của tài liệu. Xem và tải ngay bản đầy đủ của tài liệu tại đây (1.16 MB, 40 trang )
<span class='text_page_counter'>(1)</span>Lecture PowerPoint to accompany. Molecular Biology Fifth Edition. Robert F. Weaver Chapter 13 Chromatin Structure and Its Effects on Transcription Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display..
<span class='text_page_counter'>(2)</span> Chromatin Structure • Eukaryotic genes do not exist naturally as naked DNA, or even as DNA molecules bound only to transcription factors • They are complexed with an equal mass of other proteins to form chromatin • Chromatin is variable and the variations play an enormous role in chromatin structure and in the control of gene expression 13-2.
<span class='text_page_counter'>(3)</span> 13.1 Histones • Eukaryotic cells contain 5 kinds of histones – – – – –. H1 H2A H2B H3 H4. • Histone proteins are not homogenous due to: – Gene reiteration – Posttranslational modification. 13-3.
<span class='text_page_counter'>(4)</span> Properties of Histones • Abundant proteins whose mass in nuclei nearly equals that of DNA • Pronounced positive charge at neutral pH • Most are well-conserved from one species to another • Not single copy genes, repeated many times – Some copies are identical – Others are quite different – H4 has only had 2 variants ever reported 13-4.
<span class='text_page_counter'>(5)</span> 13.2 Nucleosomes • Chromosomes are long, thin molecules that will tangle if not carefully folded • Folding occurs in several ways • First order of folding is the nucleosome, which have a core of histones, around which DNA winds – X-ray diffraction has shown strong repeats of structure at 100Å intervals – This spacing approximates the nucleosome spaced at 110Å intervals 13-5.
<span class='text_page_counter'>(6)</span> Histones in the Nucleosome • Chemical cross-linking in solution: – H3 to H4 – H2A to H2B. • H3 and H4 exist as a tetramer (H3-H4)2 • Chromatin is composed of roughly equal masses of DNA and histones – Corresponds to 1 histone octamer per 200 bp of DNA – Octamer composed of: • 2 each H2A, H2B, H3, H4 • 1 each H1 13-6.
<span class='text_page_counter'>(7)</span> H1 and Chromatin • Treatment of chromatin with trypsin or high salt buffer removes histone H1 • This treatment leaves chromatin looking like “beads-on-a-string” • The beads named nucleosomes – Core histones form a ball with DNA wrapped around the outside – DNA on outside minimizes amount of DNA bending – H1 also lies on the outside of the nucleosome 13-7.
<span class='text_page_counter'>(8)</span> Nucleosome Structure • Central (H3-H4)2 core attached to H2AH2B dimers • Grooves on surface define a left-hand helical ramp – a path for DNA winding – DNA winds almost twice around the histone core condensing DNA length by 6- to 7-X – Core histones contain a histone fold: • 3 -helices linked by 2 loops • Extended tail of abut 28% of core histone mass • Tails are unstructured 13-8.
<span class='text_page_counter'>(9)</span> Crystal Structure of a Nucleosomal Core Particle. 13-9.
<span class='text_page_counter'>(10)</span> The 30-nm Fiber • Second order of chromatin folding produces a fiber 30 nm in diameter – The string of nucleosomes condenses to form the 30-nm fiber in a solution of increasing ionic strength – This condensation results in another six- to seven-fold condensation of the nucleosome itself. • Four nucleosomes condensing into the 30nm fiber form a zig-zag structure 13-10.
<span class='text_page_counter'>(11)</span> Models for the 30-nm Fiber • The solenoid and the two-start double helix model each have experimental support • A technique called single-molecule force spectroscopy was employed to answer the question, ‘which model is correct?’ • Results suggested that most of the chromatin in a cell (presumably inactive) adopts a solenoid shape while a minor fraction (potentially active) forms a 30-nm fiber according to the two-start double helix 13-11.
<span class='text_page_counter'>(12)</span> Higher Order Chromatin Folding • 30-nm fibers account for most of chromatin in a typical interphase nucleus • Further folding is required in structures such as the mitotic chromosomes • Model favored for such higher order folding is a series of radial loops. Source: Adapted from Marsden, M.P.F. and U.K. Laemmli, Metaphase chromosome structure: Evidence of a radial loop model. Cell 17:856, 13-12 1979..
<span class='text_page_counter'>(13)</span> Relaxing Supercoiling in Chromatin Loops • When histones are removed, 30-nm fibers and nucleosomes disappear • Leaves supercoiled DNA duplex • Helical turns are superhelices, not ordinary double helix • DNA is nicked to relax. 13-13.
<span class='text_page_counter'>(14)</span> 13.3 Chromatin Structure and Gene Activity • Histones, especially H1, have a repressive effect on gene activity in vitro • Histones play a predominant role as regulators of genetic activity and are not just purely structural • The regulatory functions of histones have recently been elucidated. 13-14.
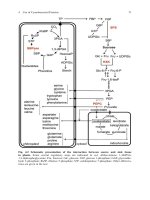
<span class='text_page_counter'>(15)</span> Effects of Histones on Transcription of Class II Genes • Core histones assemble nucleosome cores on naked DNA • Transcription of reconstituted chromatin with an average of 1 nucleosome / 200 bp DNA exhibits 75% repression relative to naked DNA • Remaining 25% is due to promoter sites not covered by nucleosome cores 13-15.
<span class='text_page_counter'>(16)</span> Histone H1 and Transcription • Histone H1 causes further repression of template activity, in addition to that of core histones • H1 repression can be counteracted by transcription factors • Sp1 and GAL4 act as both: – Antirepressors preventing histone repressions – Transcription activators. • GAGA factor: – Binds to GA-rich sequences in the Krüppel promoter – An antirepressor – preventing repression by histones 13-16.
<span class='text_page_counter'>(17)</span> A Model of Transcriptional Activation. 13-17.
<span class='text_page_counter'>(18)</span> Nucleosome Positioning • Model of activation and antirepression asserts that transcription factors can cause antirepression by: – Removing nucleosomes that obscure the promoter – Preventing initial nucleosome binding to the promoter. • Both actions are forms of nucleosome positioning – activators force nucleosomes to take up positions around, not within, promoters 13-18.
<span class='text_page_counter'>(19)</span> Nucleosome-Free Zones • Nucleosome positioning would result in nucleosome-free zones in the control regions of active genes • Assessment in SV40 DNA, a circular minichromosome, was performed to determine the existence of nucleosome-free zones - with the use of restriction sites it was found that the late control region is nucleosome free. 13-19.
<span class='text_page_counter'>(20)</span> Detecting DNase-Hypersensitive Regions • Active genes tend to have DNase-hypersensitive control regions • Part of this hypersensitivity is due to absence of nucleosomes. 13-20.
<span class='text_page_counter'>(21)</span> Histone Acetylation • Histone acetylation occurs in both cytoplasm and nucleus • Cytoplasmic acetylation carried out by HAT B (histone acetyltransferase, HAT) – Prepares histones for incorporation into nucleosomes – Acetyl groups later removed in nucleus. • Nuclear acetylation of core histone N-terminal tails – Catalyzed by HAT A – Correlates with transcription activation – Coactivators of HAT A found which may allow loosening of association between nucleosomes and gene’s control region – Attracts bromodomain proteins, essential for transcription. 13-21.
<span class='text_page_counter'>(22)</span> Histone Deacetylation • Transcription repressors bind to DNA sites and interact with corepressors which in turn bind to histone deacetylases – Repressors • Mad-Max. – Corepressors • NCoR/SMRT • SIN3. – Histone deacetylases - HDAC1 and 2 13-22.
<span class='text_page_counter'>(23)</span> Model for participation of HDAC in transcription repression • Assembly of complex brings histone deacetylases close to nucleosomes • Deacetylation of core histones allows – Histone basic tails to bind strongly to DNA, histones in neighboring nucleosomes – This inhibits transcription. 13-23.
<span class='text_page_counter'>(24)</span> Model for Activation and Repression. 13-24.
<span class='text_page_counter'>(25)</span> Chromatin Remodeling • Activation of many eukaryotic genes requires chromatin remodeling • Several protein complexes carry this out – All have ATPase harvesting energy from ATP hydrolysis for use in remodeling – Remodeling complexes are distinguished by ATPase component. 13-25.
<span class='text_page_counter'>(26)</span> Remodeling Complexes • SWI/SNF – In mammals, has BRG1 as ATPase – 9-12 BRG1-associated factors (BAFs) • A highly conserved BAF is called BAF 155 or 170 • Has a SANT domain responsible for histone binding • This helps SWI/SNF bind nucleosomes. • ISWI – Have a SANT domain – Also have SLIDE domain involved in DNA binding. 13-26.
<span class='text_page_counter'>(27)</span> Models for SWI/SNF Chromatin Remodeling. 13-27.
<span class='text_page_counter'>(28)</span> Mechanism of Chromatin Remodeling • Mechanism of chromatin remodeling involves: – Mobilization of nucleosomes – Loosening of association between DNA and core histones. • Catalyzed remodeling of nucleosomes involves formation of distinct conformations of nucleosomal DNA/core histones when contrasted with: – Uncatalyzed DNA exposure in nucleosomes – Simple nucleosome sliding along a DNA stretch 13-28.
<span class='text_page_counter'>(29)</span> Remodeling in Yeast HO Gene Activation • Chromatin immunoprecipitation (ChIP) can reveal the order of binding of factors to a gene during activation • As HO gene is activated: – First factor to bind is Swi5 – Followed by SWI/SNF and SAGA containing HAT Gcn5p – Next general transcription factors and other proteins bind. • Chromatin remodeling is among the first steps in activation of this gene • Order could be different in other genes 13-29.
<span class='text_page_counter'>(30)</span> Remodeling in the Human IFN- Gene: The Histone Code The Histone Code:. – The combination of histone modifications on a given nucleosome near a gene’s control region affects efficiency of that gene’s transcription – This code is epigenetic, not affecting the base sequence of DNA itself. • Activators in the IFN- enhanceosome can recruit a HAT (GCN5) – HAT acetylates some Lys on H3 and H4 in a nucleosome at the promoter – Protein kinase phosphorylates Ser on H3 – This permits acetylation of another Lys on H3. 13-30.
<span class='text_page_counter'>(31)</span> Remodeling in the Human IFN- Gene: TF Binding • Remodeling allows TFIID to bind 2 acetylated lysines in the nucleosome through the dual bromodomain in TAF1 • TFIID binding – Bends the DNA – Moves remodeled nucleosome aside – Paves the way for transcription to begin. 13-31.
<span class='text_page_counter'>(32)</span> Heterochromatin • Euchromatin: relatively extended and open chromatin that is potentially active • Heterochromatin: very condensed with its DNA inaccessible – Microscopically appears as clumps in higher eukaryotes – Repressive character able to silence genes as much as 3 kb away. 13-32.
<span class='text_page_counter'>(33)</span> Heterochromatin and Silencing • Formation at the tips of yeast chromosomes (telomeres) with silencing of the genes is the telomere position effect (TPE). • Depends on binding of proteins – RAP1 to telomeric DNA – Recruitment of proteins in this order: • SIR3 • SIR4 • SIR2 13-33.
<span class='text_page_counter'>(34)</span> SIR Proteins • Heterochromatin at other locations in chromosome also depends on the SIR proteins • SIR3 and SIR4 interact directly with histones H3 and H4 in nucleosomes – Acetylation of Lys 16 on H4 in nucleosomes prevents interaction with SIR3 – Blocks heterochromatin formation. • Histone acetylation also works in this way to promote gene activity 13-34.
<span class='text_page_counter'>(35)</span> Histone Methylation • Methylation of Lys 9 in N-terminal tail of H3 attracts HP1 • This recruits a histone methyltransferase – Methylates Lys 9 on a neighboring nucleosome – Propagates the repressed, heterochromatic state. • Methylation of Lys and Arg side chains in core histones can have either repressive or activating effects 13-35.
<span class='text_page_counter'>(36)</span> Histone Methylation • Methylation of Lys 4 in N-terminal tail of H3 is generally tri-methylated (H3K4Me3) and is usually associated with the 5’-end of an active gene • This modification appears to be a sign of transcription initiation • Genome-wide ChIP analysis suggests that this may also play a role in controlling gene expression by controlling the re-starting of paused RNA polymerases 13-36.
<span class='text_page_counter'>(37)</span> Summary • Histone modifications can affect gene activity by two mechanisms: • 1. By altering the way histone tails interact with DNA and with histone tails in neighboring nucleosomes, and thereby altering nucleosome cross-linking • 2. By attracting proteins that can affect chromatin structure and activity. 13-37.
<span class='text_page_counter'>(38)</span> Modification Combinations • Methylations occur in a given nucleosome in combination with other histone modifications: – Acetylations – Phosphorylations – Ubiquitylations. • Each particular combination can send a different message to the cell about activation or repression of transcription • One histone modification can also influence other, nearby modifications 13-38.
<span class='text_page_counter'>(39)</span> Nucleosomes and Transcription Elongation • An important transcription elongation facilitator is FACT (facilitates chromatin transcription) – Composed of 2 subunits: • Spt16 – Binds to H2A-H2B dimers – Has acid-rich C-terminus essential for these nucleosome remodeling activities • SSRP1 binds to H3-H4 tetramers. 13-39.
<span class='text_page_counter'>(40)</span> Nucleosomes and Transcription Elongation • FACT facilitates transcription through a nucleosome by promoting loss of at least one H2A-H2B dimer from the nucleosome • Also acts as a histone chaperone promoting readdition of H2A-H2B dimer to a nucleosome that has lost such a dimer. 13-40.
<span class='text_page_counter'>(41)</span>