Tuyển chọn, cải biến và nghiên cứu đặc điểm sinh học của chủng xạ khuẩn có khả năng kháng vi khuẩn pseudomonas aeruginosa phân lập ở việt nam
Bạn đang xem bản rút gọn của tài liệu. Xem và tải ngay bản đầy đủ của tài liệu tại đây (4.52 MB, 73 trang )
VIỆN HÀN LÂM KH&CN VIỆT NAM VIỆN SINH THÁI VÀ TÀI NGUYÊN
SINH VẬT
-----------***-----------
LUẬN VĂN CAO HỌC
Mã số chuyên ngành: 60420103
Đề tài:
Tuyển chọn, cải biến và nghiên cứu đặc điểm sinh học của chủng xạ khuẩn có
khả năng kháng vi khuẩn Pseudomonas aeruginosa phân lập ở Việt Nam
Học viên: Nguyễn Huy Hùng
Lớp: CHST _ K16
Hƣớng dẫn: TS. Nguyễn Xuân Cảnh
Hà Nội, 2014
Lời cảm ơn
Tôi xin bày tỏ lòng biết ơn sâu sắc tới TS Nguyễn Xuân Cảnh Khoa Công nghệ sinh học Học viện Nông nghiệp Hà Nội, là người thầy đã hướng cho tôi những ý tưởng khoa học, tận
tnh hướng dẫn, truyền đạt kiến thức, giúp đỡ và tạo mọi điều kiện thuận lợi cho tôi hoàn
thành bản luận án này.
Tôi xin cảm ơn tất cả các thầy cô giáo Viện Sinh thái và Tài nguyên sinh vật, Viện Hàn
lâm Khoa học và công nghệ Việt Nam đã chia sẻ, động viên, giúp tôi vượt qua mọi khó khăn để
hoàn thành tốt công việc nghiên cứu của mình.
Cuối cùng, tôi xin tỏ lòng biết ơn đến gia đình và bè bạn, những người luôn bên tôi,
động viên,góp ý và tạo điều kiện tốt nhất cho tôi trong suốt thời gian học tập
và nghiên cứu.
Tác giả
Lời cam đoan
Tôi xin cam đoan đây là công trình nghiên cứu của tôi và một số kết quả cùng cộng tác
với các đồng sự khác. Các số liệu và kết quả trình bày trong luận văn là trung thực.
Hà Nội, ngày tháng
Tác giả
năm 2014
MỤC LỤC
MỤC LỤC.................................................................................................................. i Danh mục hình
.........................................................................................................
iv
Danh
..........................................................................................................v
TỔNG
QUAN
..........................................................................................3
1.1.
GIỚI THIỆU VỀ XẠ KHUẨN ....................................................................3
1.1.1. Sự phân bố và ý nghĩa của xạ khuẩn trong tự nhiên........................................3
1.1.2. Vị trí của xạ khuẩn trong sinh giới ..................................................................4
1.1.3. Cấu tạo của xạ khuẩn .......................................................................................5
1.1.4. Đặc điểm hình thái của xạ khuẩn .....................................................................8
1.1.5. Sự hình thành bào tử xạ khuẩn ........................................................................9
1.1.6. Sinh tổng hợp chất kháng sinh .......................................................................11
1.2. PHÂN LẬP CÁC CHỦNG XẠ KHUẨN SINH TỔNG HỢP CHẤT KHÁNG
SINH ........................................................................................................................13
1.2.1. Phân lập các chủng xạ khuẩn sinh chất kháng sinh .......................................13
1.2.2. Phân loại và định tên xạ khuẩn ......................................................................14
2.1. VẬT LIỆU ........................................................................................................33
2.1.1. Chủng giống vi sinh vật .................................................................................33
2.1.2. Hóa chất .........................................................................................................33
2.1.3. Thiết bị ...........................................................................................................33
2.1.4. Môi trƣờng .....................................................................................................33
2.2. PHƢƠNG PHÁP NGHIÊN CỨU ....................................................................34
2.2.1. Bảo quản giống [3].........................................................................................34
2.2.2. Xác định đặc điểm sinh học ...........................................................................35
2.2.3. Xác định sinh khối .........................................................................................36
2.2.4. Xác định hoạt tính kháng sinh [6,19].............................................................36
2.2.5. Nghiên cứu điều kiện lên men [3, 6]..............................................................37
mục
TÀI
bảng
LIỆU
2.2.6. Chạy sắc ký giấy chất kháng sinh ..................................................................38
PHẦN 3: KẾT QUẢ VÀ THẢO LUẬN .................................................................39
3.1. Phân lập vi khuẩn Pseudomonas aeruginosa ...................................................39
3.2. Xây dựng kháng sinh đồ đối với các chủng P. aeruginosa phân lập ...............41
3.3. Phân lập và tuyển chọn các chủng xạ khuẩn có khả năng kháng vi khuẩn P.
aeruginosa................................................................................................................42
3.3.1. Phân lập xạ khuẩn ..........................................................................................42
3.3.2. Sàng lọc và tuyển chọn các chủng xạ khuẩn kháng vi khuẩn P. aeruginosa 43
3.4. Nghiên cứu các đặc điểm phân loại và đặc điểm sinh học của chủng xạ khuẩn
8.9 .............................................................................................................................45
3.4.1. Đặc điểm nuôi cấy và đặc điểm hình thái ......................................................45
3.4.1. Đặc điểm hình thái .........................................................................................46
3.4.2. Một số đặc điểm sinh hóa của chủng 8.9 .......................................................46
3.4.3. Mô tả đặc điểm phân loại...............................................................................48
3.4.4. Phân loại bằng phƣơng pháp sinh học phân tử ..............................................50
3.5. Nghiên cứu động thái lên men của chủng xạ khuẩn 8.9 ...................................52
3.6. Nghiên cứu một số tính chất của dịch kháng sinh thô ......................................54
3.6.1. Tách chiết chất kháng sinh.............................................................................54
3.6.2. Độ bền nhiệt của dịch kháng thô ...................................................................54
3.6.3. Ảnh hƣởng của pH đến độ khuếch tán của dich kháng sinh thô ...................55
3.6.4. Đặc điểm sắc kí của dịch kháng sinh thô của chủng xạ khuẩn 8.9 trong một số hệ dung
môi .........................................................................................................55
3.7. Nghiên cứu cải biến chủng xạ khuẩn 8.9 ..........................................................56
PHẦN 4. KẾT LUẬN ..............................................................................................59
TÀI LIỆU THAM KHẢO .......................................................................................60
Tài liệu tiếng Việt ....................................................................................................60
Tài liệu tiếng anh......................................................................................................60
CÁC CHỮ VIẾT TẮT TRONG KHÓA LUẬN
HSCC
Hệ sợi cơ chất HSKS
Hệ sợi khí sinh RNA
Ribonucleic acide
DNA
Deoxyribonucleic acide
RA
Cuống bào tử xoắn đơn hình móc câu RF
Cuống bào tử thẳng hay lƣợn song CSBT
Cuống sinh
bào tử
BMBT
Bề mặt bào tử CKS
Chất kháng sinh TKMX
Trực khuẩn
mủ xanh
PCR
Polymerase chain reaction
Danh mục hình
Hình 1.1. Khuẩn lạc xạ khuẩn
Hình 1.2. Bào tử xạ khuẩn
Hình 1.3. Trực khuẩn mủ xanh Pseudomonas aeruginosa
Hình 3.1. Hình thái khuẩn lạc (A) và tế bào ở độ phóng đại 11000 lần trên kính hiển vi điện
tử quết của chủng vi khuẩn Pseudomonas aeruginosa phân lập tại tại bệnh viện huyết học
0
truyền máu TW sau 02 ngày nuôi cấy ở 37 C.
Hình 3.2. Thử hoạt tính kháng sinh chủng vi khuẩn Pseudomonas aeruginosa của các chủng xạ
khuẩn bằng phƣơng pháp khối thạch
Hình 3.3. Kết quả thử hoạt tính các chủng xạ khuẩn phân lập bằng phƣơng pháp giếng thạch
với vi sinh vật kiểm định là vi khuẩn Pseudomonas aeruginosa
Hình 3.4. Khả năng sinh trƣởng và màu sắc khuẩn lạc chủng 8.9 khi nuôi cấy trên môi trƣờng
ISP-1 (A) và ISP-2 (B)
Hình 3.5. Hình thái cuống sinh bào tử và bề mặt bào tử của chủng xạ khuẩn NĐ
8.9 khi quan sát trên kính hiển vi quang học ở độ phóng đại 1000 lần (A) và kính hiển vi điện
tử quết ở độ phóng đại 7500 lần
Hình 3.6. Sự sinh trƣởng của chủng xạ khuẩn 8.9 trên môi trƣờng có các nguồn cacbon khác
nhau
Hình 3.7. Sinh trƣởng và phát triển của chủng 8.9 trên môi trƣờng ISP-6
Hình 3.8. Điện di sản phẩm PCR gene 16S rRNA của chủng 8.9
Hình 3.9. Cây phát sinh chủng loại của chủng 8.9
Hình 3.10. Động thái của quá trình lên men sinh tổng hợp chất kháng sinh của chủng xạ
khuẩn 8.9 trên môi trƣờng Gauze-1
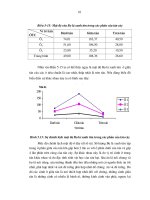
Hình 3.11. Biểu đồ giá trị Rf của chất kháng sinh thô 8.9
Danh mục bảng
Bảng 3.1.Kết quả phân lập vi khuẩn Pseudomonas aeruginosa tại Viện huyết học truyền máu
TW.
Bảng 3.2. kết quả dựng kháng sinh đồ trên máy VITEK2 compact tại bệnh viện
Huyết học truyền máu TW
Bảng 3.3. Phân loại xạ khuẩn theo nhóm màu
Bảng 3.4. Hoạt tnh kháng sinh của các chửng xạ khuẩn tuyển chọn đối với vi khuẩn
Pseudomonas aeruginosa
Bảng 3.5. Đặc điểm hình thái nuôi cấy chủng 8.9
Bảng 3.6. Khả năng sử dụng nguồn cacbon của chủng xạ khuẩn 8.9
Bảng 3.7. So sánh đặc điểm hình thái của chủng 8.9 với chủng S. parvulus Bảng 3.8.
Khả năng sử dụng nguồn cacbon của 2 chủng 8.9 và S. parvulus Bảng 3.9. So sánh
trình tự gen 16S RNA trên ngân hàng gen quốc tế
Bảng 3.10. Hoạt tnh kháng sinh của chủng xạ khuẩn 8.9 trên các môi trƣờng
nghiên cứu
Bảng 3.11. Sự biến đổi pH, hoạt tính kháng sinh, sinh khối chủng 8.9
Bảng 3.12. Hoạt tnh của dịch kháng sinh thô của chủng xạ khuẩn 8.9 đối với vi sinh vật kiểm
định Pseudomonas aruginosa ở các nhiệt độ khác nhau
Bảng 3.13. Ảnh hƣởng của pH tới sự khuếch tán của chất kháng sinh
Bảng 3.14. Giá trị Rf của dịch kháng sinh thô trên một số hệ dung môi
Bảng 3.15. Hoạt tính kháng sinh của các chủng xạ khuẩn đƣợc lựa chọn sau khi gây đột biến
đối với vi khuẩn Pseudomonas aeruginosa
MỞ ĐẦU
Pseudomonas aeruginosa (hay còn gọi là Trực khuẩn mủ xanh) là một vi khuẩn phổ
biến gây bệnh ở động vật và con ngƣời. Nó đƣợc tm thấy trong đất, nƣớc, hệ vi sinh vật
trên da và các môi trƣờng nhân tạo trên khắp thế giới. Vi khuẩn không chỉ phát triển trong
môi trƣờng không khí bình thƣờng, mà còn có thể sống trong môi trƣờng có ít khí ôxy, và do
đó có thể cƣ trú trong nhiều môi trƣờng tự nhiên và nhân tạo. Vi khuẩn này dinh dƣỡng bằng
rất nhiều các hợp chất hữu cơ; ở động vật, nhờ khả năng thích ứng vi khuẩn cho phép nó lây
nhiễm và phá hủy các mô của ngƣời bị suy giảm hệ miễn dịch. Triệu chứng chung của việc lây
nhiễm thông thƣờng là gây ra viêm nhiễm và nhiễm trùng huyết. Nếu vi khuẩn xâm nhập vào
các cơ quan thiết yếu của cơ thể nhƣ phổi, đƣờng tiết niệu, và thận, sẽ gây ra những hậu
quả chết ngƣời; vì vi khuẩn này phát triển tốt trên các bề mặt bên trong cơ thể. Vi khuẩn cũng
đƣợc phát hiện trên các dụng cụ y khoa bao gồm catheter, gây ra nhiễm khuẩn bệnh viện và
phòng mạch. Đây cũng là nguyên nhân gây ra viêm chân lông.
Nƣớc ta là một nƣớc nhiệt đới nóng ẩm thuận lợi cho vi sinh vật nói chung và xạ khuẩn
nói riêng phát triển. Do đó, có thể tm thấy nhiều chủng xạ khuẩn có khả năng sinh kháng
sinh quý, trong đó có kháng sinh kháng lại vi khuẩn Pseudomonas aruginosa gây ra các bệnh về
nhiễm trùng.
Để giải quyết những vấn đề này, chúng tôi thực hiện đề tài: “Tuyển chọn, cải biến và
nghiên cứu đặc điểm sinh học của chủng xạ khuẩn có khả năng kháng vi khuẩn Pseudomonas
aeruginosa phân lập ở Việt Nam”.
*/ Mục tiêu của đề tài:
- Phân lập đƣợc vi khuẩn Pseudomonas aeruginosa từ các mẫu thu thập tại một số
bệnh viện.
- Sàng lọc và tuyển chọn đƣợc chủng xạ khuẩn có khả năng kháng vi khuẩn
Pseudomonas aeruginosa từ bộ sƣu tập các chủng xạ khuẩn phân lập ở Việt Nam
1
- Phân loại và nghiên cứu một số đặc điểm sinh học của chủng xạ khuẩn đã tuyển chọn.
- Cải biến đƣợc chủng xạ khuẩn đã tuyển chọn để nâng cao hoạt tính kháng khuẩn
- Bƣớc đầu xác định đƣợc nhóm hoạt chất chính từ chủng xạ khuẩn đã tuyển chọn tác động lên
vi khuẩn Pseudomonas aeruginosa.
*/ Nội dung
- Phân lập vi khuẩn Pseudomonas aeruginosa từ các mẫu bệnh phẩm, nƣớc thải, đất thu thập
tại một số bệnh viện.
- Kiểm tra tnh mẫn cảm của các chủng Pseudomonas aeruginosa phân lập đƣợc với một số
loại chất kháng sinh
- Kiểm tra hoạt tính kháng sinh của các chủng xạ khuẩn đã phân lập đƣợc với vi khuẩn
Pseudomonas aeruginosa để tuyển chọn ra các chủng có hoạt tính cao.
- Phân loại và nghiên cứu các đặc điểm hình thái, nuôi cấy, sinh lý – sinh hóa của các
chủng xạ khuẩn tuyển chọn.
2
TỔNG QUAN TÀI LIỆU
1.1. GIỚI THIỆU VỀ XẠ KHUẨN
1.1.1. Sự phân bố và ý nghĩa của xạ khuẩn trong tự nhiên
Xạ khuẩn (Actinomycetales) là một nhóm vi khuẩn thật (Bacteria) phân bố rộng rãi trong
tự nhiên: trong đất, trong nƣớc ao hồ, một phần trong bùn và trong các cơ chất hữu cơ
khác, thậm chí trong cả cơ chất mà các vi khuẩn khác và nấm mốc không phát triển đƣợc [3]. Xạ
khuẩn có nhiều nhất trong lớp đất bề mặt, số lƣợng không chỉ phụ thuộc vào loại đất mà còn
phụ thuộc vào mức độ canh tác của đất và khả năng bao phủ của thực vật. Đất giầu dinh
dƣỡng và lớp đất trên bề mặt (đến 40cm) thƣờng có số lƣợng xạ khuẩn lớn. Số đơn vị sinh
khuẩn lạc (CFU- colony forming unit) xạ khuẩn trong 1g đất canh tác thƣờng đạt tới hàng
triệu mầm xạ khuẩn, đất hoang hoá chỉ có 10-100 nghìn mầm. Số lƣợng xạ khuẩn trong đất
cũng thay đổi theo thời gian trong năm [6, 28].
Xạ khuẩn cũng thƣờng sống trên các cây chết, rơm rạ, thƣờng hoại sinh nhƣng có thể kí
sinh trên thân cây, ở củ và rễ cây. Một số nghiên cứu cũng cho thấy sự có mặt của xạ khuẩn
trong trầm tch biển nhƣ các loài Actinomyces halophila, A. mortvalis và A. iraquensis,
Streptomyces nhƣ S. antbiotcus (MU106, MU107) và S. rimosus (MU114) [15,16, 33] hoặc
trong các mẫu nƣớc và bùn ao nuôi trồng thủy sản đặc biệt là ở lớp bùn, xác các hợp chất hữu
cơ lắng đọng nhiều.
Ngoài ra, pH của môi trƣờng cũng ảnh hƣởng đáng kể đến sự phân bố của xạ khuẩn,
thƣờng thì trong môi trƣờng trung tính hoặc hơi kiềm, mật độ xạ khuẩn cao hơn so với các
môi trƣờng có độ pH quá cao hoặc quá thấp. Xạ khuẩn thƣờng hoạt động ở pH cao hơn so
với nấm [6].
Xạ khuẩn đƣợc nghiên cứu một cách sâu sắc vì chúng có ý nghĩa quan trọng trong tự
nhiên: tch cực tham gia vào các quá trình chuyển hoá nhiều hợp chất trong đất, trong nƣớc,
chúng có vai trò rất quan trọng trong chu trình tuần hoàn vật chất trong tự nhiên. Chúng sử
dụng axit humic và các chất hữu cơ khó phân giải khác trong đất. Bên cạnh đó chúng còn sản
sinh nhiều sản phẩm trao đổi chất quan trọng nên đã
3
đƣợc dùng để sản xuất nhiều loại enzim nhƣ proteaza, amilaza, xenlulaza, glucoizomeraza,
vitamin và các axít hữu cơ [3]. Do có các loại enzim này nên chúng cũng có khả năng phân giải
các hợp chất bền vững nhƣ kitn, xenluloza, lignin…. Một đặc điểm quan trọng của xạ khuẩn là
khả năng sinh chất kháng sinh. 60-70% xạ khuẩn phân lập đƣợc từ đất có khả năng này. Trong
số khoảng 8000 chất kháng sinh hiện đã đƣợc biết đến trên thế giới thì trên 80% là do xạ khuẩn
[17]. Chúng sống trên rễ cây hoặc củ và gây bệnh, nhất là ở vùng đất kiềm, đất cát khi thời tết
khô hạn, làm cho củ lở loét, sần sùi ngay từ ngoài đồng ruộng. Một số xạ khuẩn (thuộc chi
Frankia) có thể tạo nốt sần trên rễ một số cây không thuộc bộ đậu và có khả năng cố định Nitơ
không khí thành dạng dễ sử dụng cho cây. Các chủng Streptomyces spp. sinh độc tố phytotoxin
ảnh hƣởng lớn tới mùa vụ, do chúng có thể tồn tại trong đất trên 10 năm [3, 6].
1.1.2. Vị trí của xạ khuẩn trong sinh giới
Trƣớc đây, vị trí phân loại của xạ khuẩn luôn là câu hỏi gây nhiều tranh luận giữa các
nhà vi sinh học do nó có những đặc điểm vừa giống vi khuẩn, vừa giống nấm. Trƣớc thế kỉ
19, xạ khuẩn đƣợc xếp vào nhóm nấm. Về sau, khi đã nghiên cứu sâu hơn ngƣời ta đã tm ra
các bằng chứng về mối liên hệ giữa xạ khuẩn và vi khuẩn
nhƣ:
- Một số xạ khuẩn nhƣ các loài thuộc chi Actinomyces và Nocardia rất giống với các
loài vi khuẩn thuộc chi Lactobacillus và Corynebacterium.
- Xạ khuẩn giống vi khuẩn ở chỗ không có nhân thật, chúng chỉ chứa nhiễm sắc chất
phân bố dọc theo các sợi hoặc các tế bào.
- Đƣờng kính của sợi xạ khuẩn và bào tử giống với vi khuẩn. Đồng thời xạ khuẩn
thƣờng không chứa vách ngăn.
- Xạ khuẩn là đích tấn công của các thực khuẩn thể giống nhƣ vi khuẩn, trong khi đó,
nấm không bị tấn công bởi thực khuẩn thể.
- Xạ khuẩn thƣờng nhạy cảm với các kháng sinh có tác dụng lên vi khuẩn nhƣng lại
thƣờng kháng với những kháng sinh tác dụng lên nấm nhƣ các polyen.
4
- Xạ khuẩn chứa chitn, chất có mặt trong sợi và bào tử của nhiều nấm, mà không
có ở vi khuẩn. Đồng thời giống nhƣ phần lớn vi khuẩn, xạ khuẩn không chứa xenluloza.
- Tƣơng tự với vi khuẩn, xạ khuẩn nhạy cảm với phản ứng axít của môi trƣờng, đặc điểm
này không có ở nấm.
- Các đặc điểm về sợi và nang bào tử lớn (sporangium) của chi Actinoplanes
cho thấy có thể chi này là cầu nối giữa vi khuẩn và các nấm bậc thấp.
Từ những đặc điểm trên (về nhân nguyên thuỷ, kích thƣớc nhỏ bé...) nên ngƣời ta đã
xếp xạ khuẩn vào nhóm vi khuẩn thật (Eubacteria).
Vị trí của nhóm vi khuẩn thật (Eubacteria) đƣợc xác định nhƣ sau: Năm 1978, Gibbbens
và Murray chia các vi khuẩn nhân nguyên thuỷ thành 4 ngành là ngành Gracilicutes (gồm các vi
khuẩn Gram âm), ngành Tenericute (gồm xạ khuẩn và các vi khuẩn
Mendosicutes
gồm
các
vi
khuẩn
không
Gram
dƣơng), ngành
chứa Peptdoglican trong thành tế bào, và
ngành Mollicutes gồm các vi khuẩn chƣa có thành tế bào.
Năm 1977 và 1980, Woese và cộng sự chia vi khuẩn nhân nguyên thuỷ thành 2 giới là
giới vi khuẩn thật (Eubacteria) (tƣơng đƣơng với 3 ngành Gracilicutes, Tenericute,
Mollicutes theo Gibbbens và Murray) và giới vi khuẩn cổ (Archaebacteria) (tƣơng
đƣơng với ngành Mendosicutes)[3].
Xạ khuẩn thuộc về nhóm vi khuẩn thật (Eubacteria), lớp Actinobacteria, bộ Actinomycetales,
bao gồm 10 dƣới bộ, 35 họ, 110 chi và 1000 loài. Hiện nay, 478 loài đã đƣợc công bố thuộc chi
Streptomyces và hơn 500 loài thuộc tất cả các chi còn lại đƣợc xếp vào nhóm xạ khuẩn hiếm.
1.1.3. Cấu tạo của xạ khuẩn
Xạ khuẩn giống nấm mốc ở chỗ có thể tạo thành hệ sợi, nhƣng lại là cơ thể đơn
bào, không có nhân thực (procariotc) và có kích thƣớc giống vi khuẩn. Cấu trúc khuẩn lạc xạ
khuẩn đƣợc phân biệt ở hƣớng sinh trƣởng trong và ngoài mặt môi trƣờng thạch tạo
thành khuẩn ty cơ chất và khuẩn ty khí sinh. Khuẩn ty cơ chất
5
(substrate mycelium) cắm sâu vào môi trƣờng để lấy nƣớc và thức ăn. Khuẩn ty cơ chất phát
triển một thời gian thì dài ra trong không khí thành khuẩn ty khí sinh (aerial mycelium). Ngƣời ta
gọi khuẩn ty khí sinh là khuẩn ty thứ cấp để phân biệt với khuẩn ty sơ cấp phát triển từ các bào
tử nảy mầm. Nhiều loại chỉ có khuẩn ty cơ chất nhƣng cũng có loại (nhƣ chi Sporichthya) lại chỉ
có khuẩn ty khí sinh. Khuẩn ty khí sinh phát triển ra ngoài không khí và thƣờng phần cuối các
khuẩn ty này biến thành cuống sinh bào tử. Loại khuẩn ty không mang bào tử gọi chung là
khuẩn ty dinh dƣỡng. Khuẩn ty xạ khuẩn mảnh hơn khuẩn ty nấm mốc, và có đƣờng kính
thay đổi trong khoảng từ
0,2-1,0µm đến 2-3µm. Đa số xạ khuẩn có khuẩn ty không có vách ngăn và không tự đứt đoạn.
Một số xạ khuẩn có bao sợi là một bao mỏng bọc bên ngoài khuẩn ty khí sinh, có thể thấy rất rõ
khi phân hoá thành bào tử [3].
* Màng tế bào xạ khuẩn khá phức tạp: gồm có thành tế bào và màng tế bào:
- Thành tế bào xạ khuẩn (CW) có dạng kết cấu lƣới, dày khoảng 10-20nm, có tác dụng
duy trì hình dáng của khuẩn ty và bảo vệ tế bào. Căn cứ vào kết cấu hoá học ngƣời ta chia thành
tế bào xạ khuẩn thành 4 nhóm:
Nhóm CW I: Có chứa L, L-DAP và Glixin (Streptomyces, Streptovertcillium, Sporichthya,
Nocardioides)
Nhóm CW II: Có chứa mezo-DAP và Glixin. Nhóm
CW III: Có chứa mezo-DAP.
Nhóm CW IV: Có chứa mezo- DAP, arbinoza và galactoza [3, 29].
Thành tế bào xạ khuẩn chủ yếu chia thành 3 lớp: lớp ngoài dày 60-120Å, khi già có
thể dày tới 150Å; lớp giữa rắn chắc dầy 50Å và lớp trong dày 50Å. Thành tế bào cấu tạo chủ
yếu từ các lớp glucopeptit gồm các gốc N-axetylglucozamin liên kết với N-axetylmuzamic bởi
liên kết 1,4-glucozit. Khi xử lý bằng lizozim các liên kết này bị phá vỡ, thành tế bào bị phá huỷ
và màng nguyên sinh chất bao bọc phần còn lại của tế bào tạo thành tế bào trần (protoplast).
Cấu trúc sợi cũng mất đi khi xử lý bằng hỗn hợp eter, clorofoc và các dung môi hoà tan lipit.
Điều đó chứng tỏ lớp ngoài
6
thành tế bào xạ khuẩn có cấu tạo bằng lipit. Thành tế bào xạ khuẩn không chứa
xenluloza hay kitn [3, 6, 13].
Đối với xạ khuẩn phân lập từ vùng ngập mặn, chúng có thành tế bào dày và độ bền cơ
học cao để có thể chống chịu với môi trƣờng khắc nghiệt bên ngoài (nồng độ muối trung bình
3-4%, pH 6,8 - 8,5 và nhiệt độ dao động từ 20-35C). Ở pH trung tnh và kiềm nhƣ ở biển, so
với xạ khuẩn trung tính, thành tế bào của xạ khuẩn chịu kiềm ngoài peptitdoglycan còn chứa
nhiều polyme axit nhƣ là axit galacturonic, axit gluconic, axit glutamic, axit asparic và axit
photphoric. Với điện tích âm tạo bởi thành phần axit không thuộc peptidoglican, bề mặt tế bào
+
+
có thể hấp thụ đƣợc ion Na và H
trong khi đẩy ion OH . Bên cạnh đó, tuy cấu trúc lớp pepttdoglycan là giống nhau ở
xạ khuẩn trung tnh và chịu kiềm nhƣng lại khác nhau về thành phần các hợp chất cấu thành, xạ
khuẩn ƣa kiềm chứa nhiều hesosamin và axit amin [25].
- Màng tế bào chất nằm dƣới lớp thành tế bào, dày 7,5-10,0 nm. Chúng có cấu trúc và
chức năng tƣơng tự nhƣ màng tế bào chất của vi khuẩn: chủ yếu gồm 2 thành phần là
photpholipit và protein; màng chứa nhiều enzym có vai trò đặc biệt quan trọng trong quá trình
vận chuyển và trao đổi chất qua màng [6].
* Thể trung gian (mezoxom) nằm ở phía trong màng tế bào chất, có hình phiến, hình
bọng hay hình ống. Công dụng của thể trung gian là làm tăng diện tch tếp xúc của màng tế bào
chất và do đó tăng cƣờng hoạt tính enzim, tăng chuyển điện tử...[3].
* Các vật thể ẩn nhập nằm trong tế bào chất của xạ khuẩn gồm có các hạt poliphotphat
(hình cầu, bắt mầu với thuốc nhuộm Soudan III), các hạt polisaccarit (bắt màu với dung dịch
Lugol) [3].
Xạ khuẩn là nhóm vi khuẩn Gram (+), “nhân” của xạ khuẩn cùng loại với nhân của “vi
khuẩn”, nhƣng khác với nhóm vi sinh vật không có nhân thực khác là tỉ lệ G+C cao (>70%)
trong khi đó ở vi khuẩn là 25 - 40%.
Một trong những đặc điểm đáng lƣu tâm của xạ khuẩn là chúng không bền vững về
di truyền và thƣờng xảy ra sự sắp xếp lại trong phân tử ADN. Điều này gây
7
ra một số hiện tƣợng kì lạ khác: tạo ra tnh đa dạng của hình thái, tnh kháng thuốc do sự xuất
hiện các dị vòng [3, 6, 23].
1.1.4. Đặc điểm hình thái của xạ khuẩn
Xạ khuẩn là nhóm vi khuẩn đặc biệt. Trên môi trƣờng đặc xạ khuẩn phát triển thành
những khuẩn lạc khô, kích thƣớc khuẩn lạc thay đổi tùy từng loại và điều kiện hoàn cảnh. Khuẩn
lạc xạ khuẩn không trơn ƣớt nhƣ ở vi khuẩn, nấm men mà thƣờng có dạng thô ráp, có các
nếp toả ra theo hình phóng xạ vì vậy mới có tên gọi là xạ khuẩn (actinomycetes, tếng Hi
Lạp: Akts là “ta”, mykes là “nấm”). Khuẩn lạc xạ khuẩn thƣờng chắc (dùng que cấy không di
đƣợc khuẩn lạc của xạ khuẩn vì khuẩn ty cơ chất bám sâu trong thạch). Mặt khuẩn lạc xù xì, có
dạng da, dạng vôi, dạng nhung tơ hay dạng màng dẻo. Khuẩn lạc của xạ khuẩn không thể lẫn
với khuẩn lạc của nấm vì khuẩn ty của xạ khuẩn thƣờng mảnh hơn của nấm mốc (khuẩn ty cơ
chất 0,8µm; khuẩn ty khí sinh 1,14µm), đƣờng kính khuẩn ty nấm có thể lớn hơn 10 lần
đƣờng kính khuẩn ty của xạ khuẩn [3].
Khuẩn lạc xạ khuẩn thƣờng có 3 lớp: lớp vỏ ngoài là các sợi bện chặt, lớp trong tƣơng đối
xốp và lớp giữa có cấu trúc tổ ong. Khuẩn ty trong mỗi lớp có hoạt tính sinh học khác nhau. Các
sản phẩm trong qúa trình trao đổi chất nhƣ: chất kháng sinh, độc tố, enzym, vitamin, axit hữu
cơ… đƣợc tch lũy trong mỗi lớp khác nhau. Khuẩn lạc xạ khuẩn có màu sắc khác nhau: đỏ, da
cam, vàng, lam, tm tùy thuộc vào loại và điều kiện ngoại cảnh. Có loại xạ khuẩn có thể tạo
sắc tố tan trong môi trƣờng nuôi cấy, thƣờng gặp các loại mang màu: xanh, tm, đỏ, da cam,
vàng, lục, nâu và đôi khi màu đen. Có sắc tố tan trong nƣớc, có sắc tố chỉ tan trong dung môi
hữu cơ [3, 6, 34].
8
Hình 1.1. Khuẩn lạc xạ khuẩn
1.1.5. Sự hình thành bào tử xạ khuẩn
Xạ khuẩn sinh sản bằng cách tạo bào tử. Khuẩn ty khí sinh phát triển ra ngoài không khí
và thƣờng phần cuối các khuẩn ty này biến thành cuống sinh bào tử. Hai dạng cuống sinh
bào tử (sporophore) và cuống sinh nang bào tử (sporangiophorres) có thể riêng rẽ, có thể phân
nhánh.
Cuống sinh bào tử (sporophore) hình thành bào tử trần (conidia hay conidiospores). Các
chuỗi bào tử trần có thể chỉ là 1 bào tử, có thể có 2 bào tử, có thể là chuỗi ngắn hoặc dài, có
thể các bào tử trần nằm trên bó sợi (synnema), tƣơng tự bó sợi của nấm.
Cuống sinh nang bào tử (sporangiophorres) mang bào tử kín (sporangio-spores)
nằm trong túi (nang) bào và bào tử di động [3].
Hình thái, kích thƣớc của cuống sinh bào tử và bào tử là một đặc điểm quan trọng
nhất trong phân loại xạ khuẩn.
Cuống sinh bào tử thƣờng có dạng thẳng (R) hay hơi cong (F), dạng xoắn lò so (kí hiệu S)
xoắn đơn giản hình móc câu (RA). Cuống sinh bào tử dạng xoắn có chiều dài và số vòng khác
nhau. Cuống sinh bào tử dạng thẳng có thể là dài hoặc ngắn với các dạng lông cứng hoặc có
thể thon lại, uốn cong hay kéo dài. Những đặc điểm này
9
rất quan trọng khi định tên xạ khuẩn. Cuống sinh bào tử xạ khuẩn hình thành bào tử
theo 3 phƣơng thức sau:
- Phƣơng thức phát triển toàn bộ: toàn bộ hay một bộ phận của khuẩn ty hình thành ra
các thành của bào tử.
- Phƣơng thức phát triển trong thành: thành bào tử sinh ra từ tầng nằm giữa màng
nguyên sinh chất và thành khuẩn ty. Trƣờng hợp này gặp ở Planomonospora.
- Phƣơng thức phát triển bào tử nội sinh thật: thành khuẩn ty không tham gia vào
quá trình hình thành bào tử. Trƣờng hợp này gặp ở Thermoactnomycetes [3].
- Hình thái bảo tử cũng là một đặc điểm quan trọng trong định tên xạ khuẩn. Bào tử
trần (conodiospore) của xạ khuẩn có thể có hình tròn, hình bầu dục, hình que, hình trụ. Bề mặt
bào tử xạ khuẩn có dạng trơn nhẵn, xù xì, có vẩy, có gai, có lông. Bào tử trần là cơ quan
sinh sản chủ yếu của xạ khuẩn. Bào tử trần đƣợc hình thành đồng thời trên tất cả chiều dài
của cuống sinh bào tử theo hai phƣơng thức khác nhau:
- Vách ngăn hình thành dần từ phía trong của màng tế bào và tến dần vào trong tạo ra
những vách ngăn không hoàn chỉnh, sau đó sợi bào tử mới phân cắt thành các bào tử trần.
- Thành tế bào và màng tế bào đồng thời xuất hiện vách ngăn tến dần vào phía trong
làm cho sợi bào tử phân cắt, đồng thời tạo thành chuỗi bào tử trần [3].
Muốn kích thích hình thành bào tử, trƣớc hết phải kích thích sự sinh trƣởng của khuẩn
ty khí sinh. Ví dụ khi nghiên cứu 6 chủng S. griseus, Kalakoutski và Agre (1973) nhận thấy
muốn kích thích khuẩn ty khí sinh nên sử dụng các môi trƣờng có bổ sung pepton, NaCl và
glixerin. Một số tác giả khác cho rằng việc bổ sung CaCO3 và CaCl2 vào môi trƣờng cũng kích
thích quá trình hình thành bào tử ở xạ khuẩn. Tuy nhiên việc bổ sung các nguyên tố vào môi
trƣờng phải đƣợc nghiên cứu kỹ và chỉ sử dụng ở nồng độ nhất định. Môi trƣờng nuôi cấy
nghèo dinh dƣỡng sẽ kích thích hình thành bào tử ở xạ khuẩn còn môi trƣờng nuôi cấy giàu
dinh dƣỡng thì quá trình sinh bào tử thƣờng bị kìm hãm. Độ ẩm và nhiệt độ cũng có ảnh hƣởng
đến sự hình thành bào tử [6].
10
Ở các chủng xạ khuẩn sinh chất kháng sinh thuộc chi Streptomyces, khả năng sinh kháng
sinh liên quan đến sự hình thành bào tử đƣợc thể hiện rất rõ. Trong nhiều trƣờng hợp khi kích
thích hình thành bào tử, hiệu suất sinh tổng hợp chất kháng sinh giảm đi. Khi nghiên cứu đột
biến và chọn lọc chủng xạ khuẩn sản xuất oxytetraxylin và streptomyxin ngƣời ta nhận thấy
nếu sự hình thành bào tử kém thì khă năng sinh tổng hợp kháng sinh lại tăng. Đây cũng là đặc
điểm cần lƣu ý khi nghiên cứu chọn lọc các chủng có hoạt tnh cao. Nhƣ vậy, sự hình thành
bào tử và khả năng sinh kháng sinh ở xạ khuẩn có mối quan hệ nghịch [16, 24].
Hình 1.2. Bào tử xạ khuẩn
1.1.6. Sinh tổng hợp chất kháng sinh
Xạ khuẩn là nhóm sinh vật có khả năng sinh kháng sinh cao, 60 - 70% xạ khuẩn
phân lập đƣợc từ đất có khả năng này. Trong số 9000 chất kháng sinh đã đƣợc biết đến trên
thế giới thì trên 80% là do xạ khuẩn sinh ra. Chi Streptomyces là chi quan trọng nhất trong
nhóm xạ khuẩn, với trên 500 loài đã đƣợc miêu tả. Chi này có nhiều loài có khả năng sinh kháng
sinh, một trong số các kháng sinh rất quan trọng là Streptomycin do S. griseus sinh ra. Khả năng
sinh chất kháng sinh là kết quả của quá trình đối kháng giữa các vi sinh vật và thúc đẩy quá trình
tiến hoá của xạ khuẩn [17].
11
Chất kháng sinh là chất trao đổi bậc 2, thông thƣờng đƣợc hình thành ở cuối pha
sinh trƣởng, trong pha cân bằng. Ở xạ khuẩn, sinh tổng hợp chất kháng sinh có quan hệ
nghịch với sự hình thành bào tử. Đó là điểm cần lƣu ý trong khi nghiên cứu sinh tổng hợp kháng
sinh từ xạ khuẩn [12].
Đối với sinh tổng hợp chất kháng sinh ở xạ khuẩn, ngƣời ta thƣờng mô tả hai pha: pha
sinh trƣởng (trophophase): khuẩn ty sơ cấp phát triển nhanh với nguyên sinh chất đồng nhất
làm giảm nguồn cacbon, nitơ và pH; pha sinh tổng hợp (idiophase): sinh trƣởng chậm lại đôi
khi xuất hiện quá trình tự tan của khuẩn ty và sinh tổng hợp chất kháng sinh mạnh [6, 16].
Cũng có tác giả chia sự phát triển của giống thành 4 hoặc 6 pha theo các đặc điểm sinh hoá tế
bào. Trong một số trƣờng hợp quá trình sinh học này lại diễn ra mạnh mẽ trong pha sinh
trƣởng nhƣ trƣờng hợp của chủng xạ khuẩn Saccharothrix sp. SA 103 phân lập từ Sahara của
Algeria, sinh kháng sinh mutactmycin PR (Anthracycline mới). Quá trình tƣơng tự cũng đã
thấy ở chủng Saccharothrix sp. SA 233 sinh dithiolopyrrolone và chủng S. clavuligerus sinh axit
clavulanic [12,30].
Mặc dù chất kháng sinh có cấu trúc khác nhau và vi sinh vật sinh ra nó cũng đa dạng
nhƣng quá trình sinh tổng hợp chúng dƣờng nhƣ theo một số con đƣờng nhất định. Các con
đƣờng này chỉ là sự cải biên các con đƣờng sinh tổng hợp của tế bào. Các vật liệu ban đầu
của của qúa trình chuyển hoá chất trao đổi bậc hai (chất kháng sinh) là những chất trao đổi sơ
cấp. Chất kháng sinh đƣợc tổng hợp từ 1, 2 hoặc 3 chất trao đổi bậc nhất, hoặc có thể là trùng
hợp các hợp chất bậc nhất rồi biến đổi chúng bằng các enzim. Mọi con đƣờng sinh tổng hợp
chất kháng sinh đều có sự tham gia của các enzim hoặc hệ thống enzim đặc biệt. Các enzim này
đƣợc mã hoá bởi các gen nằm trong những cụm (cluster) trên nhiễm sắc thể hoặc trên plasmit.
Ví dụ tổng hợp chlotetraxyclin ở S. aureofaciens có 3 hệ thống enzim (hay hệ thống gen) tham
gia: hệ thống 1 với hơn 200 gen mã cho các enzim chuyển glucoza thành axetyl-CoA; hệ
thống 2 chuyển axetyl-CoA thành malonyl-CoA; hệ thống 3 chuyển malonyl CoA
12
thành chlotetracyclin. Tổng số gen ở 3 hệ thống liên quan tới tổng hợp chlotetracyclin lên tới
2000 gen [16,31,32,35].
Các chất kháng sinh từ xạ khuẩn rất phong phú và đa dạng, nguyên nhân là 1 loại kháng
sinh có thể do nhiều loài vi sinh vật khác nhau tổng hợp ra, cũng có khi 1 loại vi sinh vật tạo ra
nhiều loại kháng sinh khác nhau [31, 32]. Ví dụ, có tới 27 chủng xạ khuẩn khác nhau có thể
tổng hợp đƣợc kháng sinh oxytetraxyclin, hay Streptomyces rimosus cùng lúc có thể tổng
hợp đƣợc oxytetracylin và rimicidin. Ngày nay số lƣợng chất kháng sinh đƣợc sản xuất từ xạ
khuẩn tăng lên đáng kể, rất nhiều trong số đó đã đƣợc sản xuất ở quy mô thƣơng mại, có ý
nghĩa lớn trong y học và mang lại nhiều lợi nhuận.
1.2. PHÂN LẬP CÁC CHỦNG XẠ KHUẨN SINH TỔNG HỢP CHẤT KHÁNG
SINH
1.2.1. Phân lập các chủng xạ khuẩn sinh chất kháng sinh
Nhu cầu về các chất kháng sinh sử dụng trong nông nghiệp hiện nay rất lớn do sự xuất
hiện của nhiều tác nhân gây bệnh mới, cùng với hiện tƣợng kháng thuốc kháng sinh ngày
càng phổ biến. Việc tuyển chọn các chủng xạ khuẩn có khả năng đối kháng các chủng vi sinh vật
gây bệnh trên cây trồng sẽ giúp hạn chế thuốc hóa học mà vẫn có tính an toàn cao [31,32,33].
Một trong những chính sách nhằm tách và tnh sạch các chất kháng sinh mới với
mong muốn là khám phá các hệ sinh thái đặc biệt, nhƣ ở tầng nƣớc sâu, vùng biển có nồng độ
muối cao hay vùng khí hậu khô cằn. Từ năm 1975 đến 1994, một số loài xạ khuẩn mới đã
đƣợc phân lập và mô tả nhƣ Actinopolyspora halophila, Actinopolyspora mortvalis và
Actinopolyspora. Trong nỗ lực tìm kiếm những chất kháng sinh mới, gần đây các nhà khoa học
đã phân lập đƣợc một chủng xạ khuẩn kí hiệu Actinopolyspora AH1 ở Alibag, một vùng biển ở
phía tây Ấn Độ. Chủng này thuộc chi Actinopolyspora, có đặc điểm ƣa mặn (sinh trƣởng ở
nồng độ muối 815%w/v) và sinh trƣởng tối ƣu ở pH 7,6. Chủng này khác với tất cả các chủng trong
chi Actinopolyspore đã đƣợc mô tả trƣớc đây nên đƣợc coi là một chủng mới [15, 20].
13
Năm 1928, Fleming tình cờ phát hiện ra nấm Penicilium sinh kháng sinh penicilin. Nhƣng ngày
nay, cũng nhƣ các loại vi sinh vật sinh chất kháng sinh khác, xạ khuẩn đƣợc tm ra theo quy
trình tuyển chọn có hệ thống và cân nhắc kỹ lƣỡng, thƣờng tuân theo sơ đồ nhất định.
Tuy nhiên, hiếm khi phân loại đƣợc một chủng từ tự nhiên có khả năng sinh kháng
sinh cao đạt yêu cầu cho sản xuất thƣơng mại, do đó công việc tuyển chọn giống luôn gắn
liền với những thành tựu của di truyền học và đặc biệt là các phƣơng pháp làm đột biến lí hoá,
kỹ thuật khuếch đại gen nhờ plasmit. Kết quả của những nghiên cứu cơ bản này đã giúp cho
việc lựa chọn những giống có hoạt lực cao trong các điều kiện nuôi cấy ổn định và hình thành
quy trình công nghệ áp dụng trong sản xuất có quy mô rộng lớn [1, 9].
1.2.2. Phân loại và định tên xạ khuẩn
Trong số xạ khuẩn đã đƣợc đặt tên thì Actinomyces Hart
là loài lâu đời nhất.
Actinomyces israeli đƣợc coi là chủng xạ khuẩn kị khí thuần chủng đầu tên (1889), vì trƣớc khi
phát triển phƣơng thức giữ giống đông khô thì khó có thể bảo quản vi sinh vật lâu dài và
nhiều chủng giống đã bị thất lạc. Do vậy không có chủng xạ khuẩn nào đƣợc tm ra sớm hơn
các chủng mà Waksman phát hiện và lƣu trong danh mục Bộ sƣu tập giống của ông (19161919). Cho đến nay, có thêm rất nhiều chủng xạ khuẩn đƣợc lƣu trong danh mục giống xạ
khuẩn [6, 14].
Ba phƣơng pháp phân loại xạ khuẩn là phƣơng pháp phân loại truyền thống, phƣơng
pháp phân loại bằng kỹ thuật phân tử và phân loại số.
a)
Phương pháp phân loại truyền thống
Krainski (1914) lần đầu tên đề ra các chỉ têu về đặc điểm sinh lí và coi đó là mấu chốt
trong nguyên tắc phân loại để sơ bộ phân loại 17 chủng thuộc chi Actinomyces. Waksman và
Curts (1916), Waksman (1919) đã đề cập đến những dạng trung gian trong mô tả phân loại của
mình và coi đặc điểm hình thái của bào tử là đặc tnh quan trọng của các cá thể. Do vậy họ đã
đƣa ra một số loài mới có ý nghĩa. Tiếp đó Millard và Burr (1926) tm ra 17 loài, Jensen (19301931) – 2 loài, Dunche (1934)
14
– 13 loài. Baldacci và cộng sự đã bắt đầu nghiên cứu xạ khuẩn từ năm 1936 và đến năm
1953 công bố một khoá phân loại chi Streptomyces dựa trên cơ sở màu sắc khuẩn ty khí sinh,
khuẩn ty cơ chất và một số đặc điểm trung gian khác nhau [11]. Năm
1943, Waksman và Henrici đã đƣa ra một số hệ thống phân loại và đến năm 1961 có sửa đổi
[36]. Trong hệ thống này, xạ khuẩn đƣợc xếp thành nhóm gồm 3 họ, chia nhỏ thành 10 chi và
mô tả chi tết hơn 250 loài thuộc chi Streptomyces. Krassilnikov là một trong những chuyên
gia nổi tếng về nghiên cứu và phát triển khoá phân loại xạ khuẩn. Từ năm 1941 đến năm 1949
ông đã phát triển 38 loài mới. Năm 1970, ông lại công bố hệ thống phân loại mới dựa trên hệ
thống đã công bố năm 1949. Trong đó xạ khuẩn đƣợc phân chia thành 6 họ, gồm 26 chi. Theo
ông Actinomyces đƣợc dùng nhƣ một tên gọi chung cho những loài thuộc chi Streptomyces mà
Waksman và Henrici đã phác hoạ năm 1943 [14].
Một số nhà nghiên cứu ngƣời Đức (Flaig và cộng sự, 1952; Flaig và Kutzner,
1954; Kuster và Geiuz, 1955) cho rằng chi Streptomyces là một nhóm lớn. Lần đầu tên họ sử
dụng kính hiển vi điện tử để nghiên cứu đặc điểm bào tử của chi này. Kutzner đã phát triển
một khoá phân loại Streptomyces dựa trên cơ sở hình dáng cuống sinh bào tử, cấu trúc
hiển vi điện tử của bào tử, sự tạo thành sắc tố tan, hoạt phổ kháng khuẩn và một số đặc điểm
sinh lí khác [18].
Năm 1957, Gauze và cộng sự đã công bố hệ thống phân loại mới. Hệ thống này dựa vào
màu sắc khuẩn ty khí sinh, khuẩn ty cơ chất, hình dạng bào tử và cuống sinh bào tử. Hệ thống
này cũng đƣợc chỉnh lý và tái bản năm 1983. Số lƣợng hệ thống phân loại xạ khuẩn ngày
một tăng trong những năm gần đây (Praceser, 1968; Kuster,
1972; Pridham, 1974; Arai; 1969; Nonomura, 1974; Szabos, 1965…). Đó là hình thức phân loại
truyền thống dựa trên đặc điểm hính thái và tnh chất nuôi cấy. Hình thức này hiện nay vẫn
đang đƣợc sử dụng phổ biến.
b)
Phương pháp phân loại sinh học phân tử
Ngày nay nhờ sự phát triển cao của kỹ thuật, phƣơng pháp phân loại phân tử đã đƣợc
sử dụng vào phân loại xạ khuẩn. Trong hệ thống phân loại xạ khuẩn hiện nay
15
ngƣời ta vẫn dùng 3 phƣơng pháp chính: lai ADN, lai ARN và phân tch rARN 16S [37, 38]. Kết
quả đƣợc biểu hiện bằng số phần trăm sự đồng nhất (homology). Hiện nay, đại đa số các nhà
khoa học đồng ý với quan niệm hai chủng đƣợc coi là hai loài riêng biệt nếu chúng giống nhau
dƣới 70% khi tến hành lai ADN. Phân tch rARN
16S để liệt kê thứ tự các nucleott đặc trƣng của cá thể và so sánh với bảng liệt kê của cơ thể
khác để xác định mức độ giống nhau của chúng. Keswani và cộng sự đã chứng minh rằng nếu
sự tƣơng đồng giữa hai trình tự rADN 16S là 98.6% thì xác suất để mức độ giống nhau trong
phép lai ADN thấp hơn 70% sẽ là 99%. Vì thế giá trị tƣơng đồng 98.6% của trình tự rADN 16S
đƣợc coi là ngƣỡng để phân biệt hai loài khác nhau. Tuy nhiên, cũng có nhiều nhà khoa học
lấy giá trị này là 98%.
Bằng phƣơng pháp phân loại này, những kiến thức về phân loại cũng có nhiều thay đổi.
Ví dụ các nhà phân loại cho rằng chi Nocardia dị nhân (heterogeneous), nên về mặt di truyền
đã xếp loài Nocardia mediterranci sinh rifamixin vào chi Streptomyces và mang tên mới là
S. mediterranci; hợp nhất hai chi Streptomyces và Streptovertcillium vào một chi Streptomyces
hoặc chuyển loài Nocardia erythropolis vào chi Mycobacterium (mang tên mới M. rodochrous)
[23].
Vì số lƣợng các loài xạ khuẩn mới đƣợc mô tả ngày càng nhiều và để việc phân loại
nhanh, chính xác về di truyền phân tử, ngƣời ta đã sử dụng phƣơng pháp phân loại số
(dùng máy vi tnh) cũng nhƣ đƣa thêm các đặc điểm phân loại và phát sinh chủng loại.
c)
Phân loại số (Numerical taxonomy)
Phân loại số đã đƣợc Williams và cộng sự (1983) sử dụng để phân loại chi
Streptomyces và các chi có quan hệ gần gũi. Phƣơng pháp này dựa trên sự đánh giá về số lƣợng
mức độ giống nhau giữa các vi sinh vật theo một số lớn các đặc điểm, chủ yếu là các đặc
điểm hình thái, sinh lý – sinh hoá. Để so sánh các chủng với nhau từng đôi một, Sneath (1973)
đề nghị tnh hệ số giống nhau S (similarity) theo phƣơng trình sau:
16