tách dòng và thiết kế vector chuyển gen mang gen mã hóa nhân tố phiên mã gmdreb3 điều khiển tính chống chịu của cây đậu tương [glycine max (l.) merrill]
Bạn đang xem bản rút gọn của tài liệu. Xem và tải ngay bản đầy đủ của tài liệu tại đây (930.76 KB, 77 trang )
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
ĐẠ I HỌ C THÁ I NGUYÊN
TRƢỜ NG ĐẠ I HỌ C SƢ PHẠ M
BNH TH MAI ANH
TCH DÕNG V THIẾT KẾ VECTOR CHUYỂN GEN MANG GEN MÃ
HÓA NHÂN TỐ PHIÊN MÃ GmDREB3 ĐIỀU KHIỂN TÍNH CHỐNG
CHU CỦA ĐẬU TƢƠNG [Glycine max (L.) Merrill]
LUẬ N VĂN THẠ C SĨ SINH HỌ C
THI NGUYÊN - 2011
1
ĐẠ I HỌ C THÁ I NGUYÊN
TRƢỜ NG ĐẠ I HỌ C SƢ PHẠ M
LUẬ N VĂN THẠ C SĨ SINH HỌ C
TCH DÕNG V THIẾT KẾ VECTOR CHUYỂN GEN MANG GEN MÃ
HÓA NHÂN TỐ PHIÊN MÃ GmDREB3 ĐIỀU KHIỂN TÍNH CHỐNG CHU
CỦA ĐẬU TƢƠNG [Glycine max (L.) Merrill]
Chuyên ngà nh: Di truyề n họ c
M s: 62.42.70
Hc viên: Bnh Th Mai Anh
Ngườ i hướ ng dẫ n: PGS.TS. Chu Hoà ng Hà
THI NGUYÊN - 2011
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
LỜI CAM ĐOAN
Tôi xin cam đoan đây là công trình nghiên cứu của riêng tôi. Các số liệu, kết
quả nghiên cứu trong luận văn là trung thực và chƣa có ai công bố trong một công
trình nào khác. Mi trch dn đu ghi r ngun gốc.
LỜI CẢM ƠN
Tôi xin bày tỏ lòng biết ơn tới PGS.TS. Chu Hoàng Hà đã tận tình hƣớng dn, chỉ
bảo và tạo điu kiện thuận lợi cho tôi trong suốt quá trình hc tập, nghiên cứu và hoàn
thành luận văn.
Tôi xin chân thành cảm ơn TS. Lê Văn Sơn , TS. Phạm Bch Ngc, Ths. Lê Thu
Ngc cùng toàn thể cán bộ phòng Công nghệ Tế bào thực vật , Viện Công nghệ Sinh hc
đã tận tình giúp đỡ cho tôi trong quá trình nghiên cứu và hoàn thành luận văn.
Tôi xin cảm ơn các thầy cô khoa Sinh-KTNN và khoa Sau đại hc, trƣờng Đại hc
Sƣ phạm – Đại hc Thái Nguyên. Tôi xin chân thành cảm ơn tập thể cán bộ Bộ môn Di
truyn & Sinh hc hiện đại đã giúp đỡ và tạo điu kiện thuận lợi cho tôi trong suốt quá
trình hc tập và nghiên cứu đ tà i.
Tôi xin cảm ơn gia đình và bạn bè đã luôn động viên, giúp đỡ tôi trong suốt quá
trình hc tập và thực hiện thành công luận văn này.
Thái Nguyên, ngày 29 tháng 9 năm 2011
Hc viên
BNH TH MAI ANH
i
MC LC
Trang
Lờ i cam đoan
Lờ i cả m ơn
Mc lc
i
Danh mụ c chƣ̃ viế t tắ t
iv
Danh mụ c bảng
vi
Danh mụ c hình
vii
MỞ ĐẦU……………………………………………………………………………
1
I. Đt vấn đ
1
II. Mc tiêu nghiên cứu………………………………………………………
2
III. Nộ i dung nghiên cƣ́ u…………………………………………………………
2
Chƣơng 1. TỔ NG QUAN TÀ I LIỆ U……………………………………………
3
1.1. TỔNG QUAN VỀ CÂY ĐẬU TƢƠNG……………………………………
3
1.1.1. Ngun gốc phân loại cây đậu tƣơng………………………………………….
3
1.1.2. Đc điểm sinh hc của cây đậu tƣơng…………………
3
1.1.3. Giá trị kinh tế của cây đậu tƣơng
5
1.1.4. Tình hình sản xuất đậu tƣơng trên thế giới và ở Việt Nam
6
1.2. HẠN V TC ĐỘNG CỦA HẠN ĐẾN CÂY ĐẬ U TƢƠNG………
7
1.3. CC NHÓM GEN LIÊN QUAN ĐẾN TÍNH CHU HẠN…………………
11
1.3.1. Các nhóm gen liên quan đến tnh chịu hạn……………………………
11
1.3.2. Các nhân tố phiên mã điu khiển các gen liên quan đến tnh chịu hạn………
12
ii
1.3.3. Promoter rd29A……………………………………………………………
15
1.4. NHÂN TỐ ĐIỀ U KHIỂ N PHIÊN MÃ DREB……………………………
18
Chƣơng 2. VẬT LIỆU VÀ PHƢƠNG PHÁ P NGHIÊN CƢ́ U
24
2.1. VẬT LIỆU, HÓA CHẤT, THIẾT B
24
2.2. PHƢƠNG PHP NGHIÊN CỨU
25
2.2.1. Phƣơng pháp sinh học phân tử
25
2.2.1.1. Phương pháp tách chiết, tinh sạch DNA và RNA…………………………
25
2.2.1.2. Phương pháp tổng hợp cDNA………………………………………………
26
2.2.1.3. Phản ứng PCR ……………………………………
27
2.2.1.4. Phương pháp tách dòng gen………………………………………………
27
2.2.1.5. Thiết kế cấu trúc pBI101:: rd29A :: DREB3……………………………….
29
2.2.2. Phƣơng phá p tá i sinh và chuyển gen thông qua Agrobacterium
30
2.2.2.1. Phương phá p chuyể n gen vào cây đu tương thông qua Agrobacterium
30
2.2.2.2. Phương phá p chuyể n gen và o cây thuốc lá thông qua Agrobacterium
32
2.2.3. Phƣơng phá p phân tí ch cây chuyể n gen
33
2.2.3.1. Phương phá p PCR
33
2.2.3.2. Phương phá p gây hạ n nhân tạ o
33
2.2.4. Phƣơng pháp tính toán và xử lý s liệu
33
2.2.5. Phƣơng phá p xƣ̉ lý trình tƣ̣ gen và amino acid
33
Chƣơng 3. KẾT QUẢ VÀ THẢ O LUẬ N…………………………………
34
3.1. KẾ T QUẢ TCH DÕNG GEN DREB3……………………………………
34
iii
3.1.1. Kế t quả nhân gen DREB3 tƣ̀ RNA tổ ng số giố ng đậ u tƣơng Nâu Cao Bằ ng
34
3.1.2. Kế t quả ghé p nố i đoạ n gen DREB3 vào vector tách dòng …………………
35
3.1.3. Phân tích và xế p nhó m protein DREB3 dƣ̣ a và o vù ng bả o thủ AP2………….
39
3.2. KẾ T QUẢ THIẾ T KẾ VECTOR CHUYỂ N GEN DREB3 ………………
42
3.2.1. Thiế t kế vector ……………………………………………………………….
42
3.2.2.Biế n nạ p cấ u trú c vector pBI 101::rd29A::DREB3 vào vi khun
Agrobacterium tumefaciens…………………………………………
47
3.3. KẾT QUẢ CHUYỂ N GEN DREB3 VO CÂY THUỐC L THÔNG
QUA AGROBACTERIUM………………………………………………………
48
3.3.1. Kế t quả chuyể n cấ u trú c vector pBI ::rd29A::DREB3 vào ging thuc lá
K326………………………………………………………………………………….
48
3.3.2. Kế t quả phân tích và đá nh giá cá c dò ng thuố c lá thu đƣợ c……………….
50
3.4. KẾT QUẢ CHUYỂ N PROMOTER RD29A TRONG CÂY ĐẬ U
TƢƠNG……………………………………………………………………………
52
3.4.1. Kết quả chuyển cấu trúc promoter rd29A vào đậu tƣơng……………………
52
3.4.2. Kết quả phân tch cây đậu tƣơng chuyển gen bng k thuật PCR……
55
KẾ T LUẬ N VÀ ĐỀ NGHỊ …………………………………………………
56
DANH MỤ C CÔNG TRÌ NH LIÊN QUAN ĐẾ N LUẬ N VĂN…………………
57
TI LIỆU THAM KHẢO…………………………………………………………
58
PH LC…………………………………………………………………………
iv
DANH MC CHỮ CI VIẾT TẮT
ABA
Abscisic acid
ABR
ABA/stress responsive
ABRE
ABA responsive element
AP2/EREBP
APETALA2/Ethylene Response Elements Binding Proteins
AP2/ERF
APETALA2/ethylen response factor
AS
Acetosyringone
A. tumefaciens
Agrobacterium tumefaciens
BAP
6-benzyladenine
bar
gen mã hóa cho enzyme phosphinothricin acetyl transferase
bp
Base pair
bZIP
Basic leucine zipper
cDNA
Sợi DNA bổ sung (Complementary DNA)
CCM
Cocultivation medium – môi trƣờng đng nuôi cấy
Cs
Cộ ng sƣ̣
DNA
Deoxyribonucleic acid
dNTP
deoxynucleoside triphosphate
DRE/CRT
Dehydration responsive element/C repeat
DREB
Dehydration responsive element binding
EDTA
Ethylene diamine tetraacetic acid
E. coli
Escherichia coli
GA
3
Gibberellic acid
GM
Germination medium – môi trƣờng nảy mầm hạt
v
Gus
β –Glucuronidase gene (Gen mã hóa enzyme β-Glucuronidase)
IBA
Indole-3-butyric acid
IPTG
Isopropylthio-beta-D-galactoside
Kb
kilo base
LB
Luria Bertani
LEA
Late embryogenesis abundant
MAP kinase
Mitogen-activated protein kinases
MES
2-[N-morpholino] ethanesulfonic acid
MS
Môi trƣờng muối cơ bản theo Murashige và Skoog (1962)
NSL
Nuclear localization signal
OD
Optical density
rd29A
Responsive Dehydration
PCR
Polymerase Chain Reaction – phản ứng chuỗi polymerase
RM
Rooting medium – môi trƣờng ra rễ
RNA
Ribonucleic acid
SDS
Sodium dodecylsulfat
SIM
Shoot induction medium – môi trƣờng tạo đa chi
SEM
Shoot elongation medium – môi trƣờng kéo dài chi
TBE
Tris - Boric acid – EDTA
TAE
Tris - Acetate – EDTA
TE
Tris – EDTA
v/p
vòng/phút
X-gal
5-brom- 4-chloro-3-indolyl-b-D-galactosidase
vi
DANH MỤ C BẢ NG
Trang
Bảng 1.1. Tình hình sản xuất đậu tƣơng ở Việt Nam từ năm 2003 đến 2010…
6
Bảng 2.1. Thành phần dung dịch đệm tách chiết……………………………
25
Bảng 2.2. Thành phần của phản ứng PCR…………………………………….
27
Bảng 2.3. Thành phần phản ứng gắn gen vào vector tách dòng……………….
27
Bảng 2.4. Thành phần phản ứng colony – PCR……………………………….
28
Bảng 2.5. Thành phần hoá chất tách plasmid………………………………….
29
Bảng 2.6. Thành phần phản ứng cắt bằ ng enzyme BamHI, SacI
29
Bảng 2.7. Môi trƣờ ng nuôi cấ y đậ u tƣơng…………………………………….
31
Bảng 2.8. Môi trƣờ ng nuôi cấ y thuố c lá ……………………………………….
32
Bảng 3.1. Sƣ̣ sai khá c về trình tƣ̣ nucleotide và amino acid củ a 3 trình tự gen
DREB3 thu đƣợ c vớ i gen GmDREB3 mã số DQ055133………………………
38
Bảng 3.2. Tỷ lệ số ng só t của của các mảnh thuốc lá biến nạp qua các giai
đoạn chọ n lọ c
48
Bảng 3.3. Tỷ lệ số ng só t của các mô đậ u tƣơng qua các giai đoạn chọ n lọ c
53
vii
DANH MỤ C HÌ NH
Trang
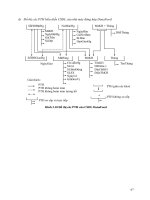
Hnh 1.1. Hệ thống nhân tố phiên mã điề u khiể n sƣ̣ biể u hiệ n cá c gen chƣ́ c năng
tham gia phả n ƣ́ ng chố ng chịu vớ i điề u kiệ n bấ t lợ i củ a Arabidopsis …………
14
Hnh 1.2. Sơ đồ về sƣ̣ trả lờ i cá c tá c nhân stress môi trƣờ ng ……………………
17
Hnh 3.1. Kế t quả nhân gen DREB3 ………………………………………………
34
Hnh 3.2. Kế t quả biế n nạ p plasmid tá i tổ hợ p và o tế bà o khả biến DH5α ……
35
Hnh 3.3. Hình ảnh điện di sản phm colony-PCR chọ n lọ c khuẩ n lạ c mang gen
DREB3 …………………………………………………………………………
36
Hnh 3.4. Điệ n di sả n phẩ m cắ t kiể m tra plamid bằ ng enzyme BamHI …………
36
Hnh 3.5. Kế t quả so sánh trình tự nucleotide …………………………………
37
Hnh 3.6. Kế t quả phân tích trình tƣ̣ amino acid củ a protein DREB3 …………
39
Hnh 3.7. Kế t quả phân tí ch trình tƣ̣ amino acid suy diễ n cá c gen họ AP 2/ERF
trên phầ n mề m Clustal W ………………………………………………………
40
Hnh 3.8. Phân tích cây phả hệ cá c gen họ AP2/ERF sƣ̉ dụ ng phầ n mề m Cobalt
41
Hnh 3.9. Mô hình mô tả cá c bƣớ c thiế t kế vector chuyể n gen …………………
42
Hnh 3.10. Kế t quả điệ n di sả n phẩ m PCR vớ i cặ p mồ i DREB3-BamHI/DREB3-
SacI ………………………………………………………………………………
43
Hnh 3.11. Kế t quả điệ n di sả n phẩ m phả n ƣ́ ng cắ t pBI::rd29A::gus bằ ng BamHI
và SacI
44
Hnh 3.12. Hình ảnh điện di sản phm colony-PCR vớ i cặ p mồ i đặ c hiệ u ……
45
Hnh 3.13. Hình ảnh điện di sản phm cắt kiểm tra plasmid ……………………
46
Hnh 3.14. Kế t quả điệ n di sả n phẩ m colony-PCR vớ i cặ p mồ i đặ c hiệ u ………
47
Hnh 3.15. Các giai đoạn phát triển của mảnh thuốc lá biến nạp …………………
49
Hnh 3.16. Kế t quả điệ n di cá c sả n phẩ m PCR cá c dò ng thuố c lá thu đƣợ c ……
50
Hnh 3.17. Kế t quả gây hạ n nhân tạ o cá c dò ng thuố c lá chuyể n
gen………………………………………………………………………………….
51
Hnh 3.18. Các giai đoạn của quá trình chuyển gen ở cây đậ u tƣơng …………
54
Hnh 3.19. Kế t quả điệ n di sả n phẩ m PCR vớ i hai cặ p mồ i đặ c hiệ u củ a promoter
rd29A và cp mi nhân gen virC…………………………………………………
55
1
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
MỞ ĐẦU
I. Đt vấn đề
Đậu tƣơng (Glycine max (L.) Merrill) là cây trng chnh ở Việt Nam chỉ xếp sau
cây lúa và ngô, là ngun nguyên liệu chế biến thức ăn gia súc và cung cấp ngun đạm
thực vật cho ngƣời. Đậu tƣơng là cây thực phm ngắn ngày có giá trị kinh tế cao , có tá c
dng trong cải tạo đất trng , dễ canh tác và đc biệt có khả năng thch nghi với nhiu
vùng sinh thái khác nhau. Vì vậy, cây đậu tƣơng đã đƣợc trng ở trên 200 quốc gia, với
sản lƣợng trung bình hàng năm khoảng hơn 250 triệu tấn [41]. Từ vị tr là một nƣớc xuất
khu đậu tƣơng vào những năm 1980, đến nay Việt Nam đã trở thành một nƣớc nhập
khu đậu tƣơng với số lƣợng lớn, riêng năm 2010 là 227,651 tấ n [43]. Vì vậy, việ c phá t
triể n diệ n tích và sả n lƣợ ng đậ u tƣơng là mộ t chiế n lƣợ c quan trọ ng trong chiế n lƣợ c phá t
triể n nông nghiệ p ở Việ t Nam hiệ n nay.
Các yếu tố bất lợi của môi trƣờng hiện đang là những thách thức lớn cho mc tiêu
duy trì sƣ̣ phá t triể n bề n vƣ̃ ng trong sả n xuấ t lƣơng thƣ̣ c cho con n gƣờ i, trong đó hạn và
mặ n là hai trong số nhƣ̃ ng nhân tố quan trọ ng nhấ t kì m hã m sƣ̣ phá t triể n sản xuất nông
nghiệ p. Những năm gần đây, thế giới cũng nhƣ Việt Nam thƣờng xuyên phải gánh chịu
những biến động lớn; sự gia tăng nhiệt độ trái đất, hạn hán, lũ lt, xói mòn, thoái hóa đất
gây ảnh hƣởng không nhỏ tới năng suất, sản lƣợng, chất lƣợng cây trng.
Đậu tƣơng là cây chịu hạn kém [3], [10], khi thiếu nƣớc ở các thời kỳ khác đu có
ảnh hƣởng xấu đến năng suất. Nhƣ vậy, việc chn và tạo đƣợc các giống đậu tƣơng có
khả năng chống chịu hạn là một nhu cầu cần thiết trong sản xuất và đƣợc xem nhƣ định
hƣớng nghiên cứu phát triển cây đậu tƣơng ở Việt Nam.
Cách tiếp cậ n hƣớ ng nghiên cƣ́ u về stress thƣ̣ c vậ t hiệ n nay đang tậ p trung và o
phân lậ p và nghiên cƣ́ u đặ c tính mộ t tậ p hợ p gen liên quan đế n tá c độ ng bấ t lợ i củ a môi
trƣờ ng và mố i liên hệ vớ i cá c stress mặ n , hạn và nhiệt độ . Hàng trăm gen đƣợ c cả m ƣ́ ng
trong cá c điề u kiệ n bấ t lợ i khá c nhau và sả n phẩ m củ a cá c gen cả m ƣ́ ng vớ i điề u kiệ n bấ t
lợ i đƣợ c chia là m hai nhó m : (1) nhóm các protein giúp thực vật chống lại bất lợi của môi
trƣờ ng; (2) nhóm các prot ein là m nhiệ m vụ điề u hò a biể u hiệ n gen và truyề n tín hiệ u
2
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
trong quá trình đá p ƣ́ ng điề u kiệ n bấ t lợ i , nhóm này chnh là cá c nhân tố phiên mã . Gầ n
đây, dƣ̣ a trên nhƣ̃ ng thà nh tƣ̣ u củ a chƣơng trình nghiên cƣ́ u chƣ́ c năng hệ gen thƣ̣ c vậ t
cùng với sự phát triển các k thuật microarray , proteomic…, các nhà khoa hc đã phát
hiệ n và chƣ́ ng minh vai trò quan trọ ng củ a cá c nhó m gen điề u khiể n (Transcription factor,
Kinase) trong việ c tăng cƣờ ng tí nh chịu hạ n ở th ực vậ t . Nhóm gen điu khiển mc dù
không tham gia trƣ̣ c tiế p và o phả n ƣ́ ng đá p ƣ́ ng vớ i điề u kiệ n hạ n củ a thƣ̣ c vậ t , nhƣng sƣ̣
biể u hiệ n củ a chú ng lạ i có vai trò kí ch hoạ t sƣ̣ biể u hiệ n củ a rấ t nhiề u gen chƣ́ c năng khá c
tham gia và o quá trình đá p ƣ́ ng hạ n, dẫ n tớ i là m tăng cƣờ ng khả năng chịu hạ n ở thƣ̣ c vậ t.
Phát hiện này đã mở ra một hƣớng nghiên cứu mới cho lnh vực chn giống chuyển gen ở
thƣ̣ c vậ t, đó là chỉ chuyể n mộ t hay và i gen điề u khiể n, thay vì và i trăm gen chƣ́ c năng và o
cây để tăng cƣờ ng tí nh chố ng chịu củ a cây trồ ng [8].
Xuất phát từ nhƣ̃ ng cơ sở trên, chúng tôi tiến hành đ tài: “Tách dng và thiế t kế
vector chuyể n gen mang gen mã hó a nhân tố phiên m GmDREB3 điề u khiể n tính
chố ng chịu củ a cây đậ u tƣơng [Glycine Max (L.) Merrill]”.
II. Mc tiêu nghiên cứu
- Phân lập đƣợc cDNA của gen DREB3 tƣ̀ cây đậu tƣơng.
- Thiết kế đƣợc vector chuyển gen mang cấu trúc rd29A::GmDREB3.
- Đánh giá hoạt động của cấu trúc rd29A::GmDREB3 trên cây thuốc lá chuyển gen.
- Kiểm tra khả năng hoạt động của promoter rd29A trên cây đậu tƣơng chuyển gen.
III. Nộ i dung nghiên cƣ́ u
- Tách dòng, xác định và phân tch trình tự cDNA của gen DREB3.
- Thiết kế vector chuyển gen mang cấu trúc rd29A::GmDREB3.
- Tạo cây thuốc lá chuyển gen mang cấu trúc rd29A::GmDREB3 và đánh giá khả
năng chịu hạn của cây thuốc lá chuyển gen tạo đƣợc.
- Tạo cây đậu tƣơng chuyển gen mang cấu trúc pBI::rd29A::gus.
3
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
Chƣơng 1
TỔ NG QUAN TÀ I LIỆ U
1.1.TỔNG QUAN VỀ CÂY ĐẬU TƢƠNG
1.1.1. Nguồn gc phân loại cây đậu tƣơng
Đậu tƣơng là một trong những loài cây trng đƣợc biết đến từ rất sớm. Các bng
chứng v lịch sử, địa lý và khảo cổ hc đu chỉ ra rng đậu tƣơng có nguyên sản ở châu Á
và có ngun gốc từ Trung Quốc. Đậu tƣơng đƣợc thuần hóa và trng làm cây thực phm ở
Trung Quốc vào khoảng thế kỉ 17 trƣớc công nguyên. Sau đó, nó đƣợc truyn bá sang
Nhật Bản vào khoảng thế kỷ thứ 8, du nhập vào nhiu nƣớc châu Á khác nhƣ Indonesia,
Philippin, Thái Lan, Ấn Độ, Việt Nam,… vài thế kỷ sau đó. Đậu tƣơng đƣợc trng ở châu
Âu vào thế kỷ 17 và ở Hoa Kỳ thế kỷ 18. Và ngày nay, Hoa Kỳ là quốc gia sản xuất đậu
tƣơng hàng đầu thế giới, chiếm 50% sản lƣợng trên toàn thế giới, tiếp theo là các nƣớc
Trung Quốc, Ấn Độ…[41]
Cây đậu tƣơng thuộc bộ Phaseoleae, h Đậu Fabaceae, h ph cánh bƣớm
Papilionoideae, chi Glycine. Đậu tƣơng có tên khoa hc là Glycine Max (L) Merr. Có
nhiu hệ thống phân loại khác nhau, trong đó hệ thống phân loại căn cứ vào đc điểm
hình thái, sự phân bố địa lý và số lƣợng nhiễm sắc thể do Hymowit và Newell (1984) xây
dựng đƣợc công nhận rộng rãi. Theo cách phân loại này thì ngoài chi Glycine còn có chi
ph Soja. Chi Glycine đƣợc chia thành 7 loài hoang dại lâu năm, chi Soja gm 2 loài
trong đó có loài đậu tƣơng trng Glycine Max (L) Merr [3].
1.1.2. Đc điểm sinh học của cây đậu tƣơng
Đậu tƣơng (Glycine max (L.) Merril) là cây hai lá mầm, thân thảo. Thân cây có
nhiu lông nhỏ, khi còn non thân có màu xanh hoc tm, khi v già chuyển màu nâu nhạt.
Màu sắc của thân cây cho ta biết màu của hoa sau này. Nếu thân có màu xanh, hoa sẽ có
màu trắng, nếu thân có màu tm thì sau hoa sẽ có màu tm đỏ. Chiu cao của thân từ 0,3 –
1m tùy theo giống. Khác với những cây trng khác thân cây đậu tƣơng phát triển mạnh
nhất lại chnh vào lúc cây ra hoa rộ nhất. Đây là lúc mà giai đoạn sinh trƣởng sinh dƣỡng
4
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
và sinh trƣởng sinh thực cạnh tranh nhau dn đến khủng hoảng dinh dƣỡng. Vì vậy, trng trt
cần chú ý cung cấp đầy đủ dinh dƣỡng cho cây trƣớc khi chúng bƣớc vào giai đoạn này [3].
Cây đậu tƣơng có ba loại lá: lá mầm, lá đơn, lá kép. Mỗi lá kép thƣờng có 3 lá chét.
Lá kép mc so le, trên phiến lá có nhiu lông tơ. Hình dạng của lá thay đổi theo giống,
những giống có lá dài và nhỏ chịu hạn tốt nhƣng cho năng suất thấp. Những giống lá to
thƣờng cho năng suất cao hơn nhƣng cũng chịu hạn kém hơn [3].
Hoa đậu tƣơng đƣợc phát sinh từ nách lá, đầu cành hoc đầu thân. Hoa đậu tƣơng
mc thành chùm, mỗi chùm thƣờng có từ 3-5 hoa. Hoa lƣỡng tnh nên đậu tƣơng là cây tự
th phấn, tỷ lệ giao phấn rất thấp chỉ chiếm 0,5-1%. Thời gian bắt đầu ra hoa sớm hay
muộn, dài hay ngắn ph thuộc vào từng giống đậu tƣơng. Căn cứ vào phƣơng thức ra hoa
ngƣời ta chia đậu tƣơng ra làm hai nhóm: nhóm ra hoa hữu hạn, hƣớng ra hoa từ trên
xuống dƣới và từ ngoài vào trong và nhóm ra hoa vô hạn, hƣớng ra hoa từ dƣới lên trên
và từ trong ra ngoài [3].
Quả đậu tƣơng thẳng hoc hơi cong, dài từ 2-7cm. Mỗi quả thƣờng có 2-3 hạt. Hạt
đậu tƣơng có hình tròn, bầu dc,… giống có hạt màu vàng có giá trị thƣơng phm cao.
Khối lƣợng 1000 hạt dao động trung bình từ 100-200g. Hình dạng và màu sắc của rốn hạt
đc trƣng cho mỗi giống [3].
Bộ rễ đậu tƣơng gm rễ chnh và rễ ph. Trên rễ có rất nhiu nốt sần, đó là kết quả
của sự cộng sinh giữa vi khun Rhizobium japonicum với rễ. Nốt sần có thể dài 1 cm,
đƣờng knh 5-6 mm, khi mới hình thành nó có màu trắng sữa, khi phát triển tốt nhất nốt
sần có màu hng. Nốt sần tập trung nhiu ở tầng đất có độ sâu từ 0-20cm, có vai trò quan
trng trong việc cố định đạm từ nitơ không kh, với lƣợng đạm cung cấp cho cây khoảng
30-60kg/ha [3].
Dựa vào thời gian sinh trƣởng, đậu tƣơng đƣợc chia thành các loại: chn rất sớm:
thu hoạch sau 80-90 ngày, chn sớm thu hoạch sau 90-100 ngày, chín trung bình thu
hoạch sau 100-110 ngày, chn muộn trung bình cho thu hoạch sau 110-120 ngày, chín
muộn cho thu hoạch sau 130-140 ngày, chn rất muộn cho thu hoạch sau 140-150 ngày.
5
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
Thời gian sinh trƣởng của mỗi giống đƣợc đánh giá theo từng vùng và v trng nhất định do
thời gian sinh trƣởng của nó chịu ảnh hƣởng nhiu bởi thời gian chiếu sáng và nhiệt độ [3].
1.1.3. Giá trị kinh tế của cây đậu tƣơng
Đậu tƣơng là cây trng ngắn ngày có giá trị kinh tế cao, sản phm của nó đƣợc
dùng làm thực phm cho con ngƣời, thức ăn cho gia súc, nguyên liệu cho công nghiệp,
làm hàng xuất khu và là loài cây cải tạo đất trng rất tốt.
Giá trị về mt thực phẩm: hàm lƣợng protein cao cũng nhƣ nhiu hợp chất có giá
trị có trong hạt đậu khiến nó trở thành một trong những loại thực phm quan trng nhất
trên thế giới. Hàm lƣợng protein trong đậu tƣơng cao hơn trong cá, thịt và cao gấp 2 lần
các loại đậu đỗ khác, protein trong hạt đậu chiếm khoảng 35-50% (ph thuộc vào giống
và điu kiện chăm sóc), dễ tiêu hóa hơn thịt và không có thành phần tạo cholesterol. Đậu
tƣơng còn đƣợc xem là cây cung cấp dầu thực vật quan trng, lipid đậu tƣơng chứa thành
phần các acid béo không no cao, tổng số chất béo chiếm khoảng 18%. Ngoài ra trong hạt
đậu còn chứa sắt, canxi, phospho và các thành phần chất xơ tốt cho tiêu hóa. Vitamin
trong đậu tƣơng có nhiu nhóm B, đáng kể là vitamin B1, B2, B6, ngoài ra còn có vitamin
E, pholic acid. Thành phần có trong đậu tƣơng đƣợc nhắc tới nhiu và giúp ch cho sức
khỏe con ngƣời gm có phytosterol, lecithin, isoflavon và phytoestogen và những sản
phm ức chế phân hủy protein [3].
Giá trị về mt nông nghiệp: toàn bộ cây đậu tƣơng cả khi tƣơi và khô đu có thể
dùng làm thức ăn cho gia súc, sản phm ph công nghiệp nhƣ khô dầu có thành phần dinh
dƣỡng khá cao: N 6,2%; P
2
O
5
0,7%; K
2
O 2,4% là ngun thức ăn rất tốt cho gia súc. Một
kilogram hạt đậu tƣơng tƣơng đƣơng với 1,38 đơn vị thức ăn chăn nuôi. Đậu tƣơng còn
có vai trò quan trng trong cải tạo đất trng. Một hecta đậu tƣơng nếu sinh trƣởng và phát
triển tốt sẽ để lại trong đất 30-60kg N. Thân, lá với hàm lƣợng đạm cao có thể dùng để
bón cho đất thay cho phân hữu cơ. Việc luân canh đậu tƣơng với cây trng khác có thể
xem là biện pháp cải tạo đất trng hữu hiệu [3].
6
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
1.1.4. Tnh hnh sản xuất đậu tƣơng trên thế giới và ở Việt Nam
Với những giá trị kinh tế to lớn mà cây đậu tƣơng mang lại, nó đƣợc xem là một
trong những loại cây trng quan trng bậc nhất trên thế giới, chỉ đứng sau lúa mì, lúa
nƣớc và ngô.
Do có khả năng thch nghi với nhiu điu kiện kh hậu khác nhau nên đậu tƣơng
đƣợc trng rộng rãi khắp năm châu lc , tập trung nhiu nhất ở châu M tiếp đến là châu
Á. Diện tch gieo trng cũng nhƣ sản lƣợng đậu tƣơng liên tc tăng trong những năm qua.
Bình quân hàng năm trên thế giới có khoảng 91 triệu ha đậu tƣơng đƣợc gieo trng với
năng suất bình quân khá cao 22-23 tấn/ha. M là nƣớc có diện tch gieo trng cũng nhƣ
sản lƣợng đậu tƣơng lớn nhất thế giới, tiếp theo là Brazil, Achentina, Trung Quốc [41].
Ở Việt Nam, đậu tƣơng đƣợc gieo trng từ rất sớm, nó đƣợc trng trƣớc cây đậu
xanh và đậu đen. Tuy nhiên với các phƣơng pháp canh tác truyn thống, bộ giống năng
suất thấp, sản xuất nhỏ lẻ, giá thành cao, lãi suất thấp, giá thành đậu tƣơng trong nƣớc
không có khả năng cạnh tranh với đậu tƣơng nhập khu là nguyên nhân khiến cho nông
dân không mn mà với cây đậu tƣơng. Vì vậy, diện tch và sản lƣợng đậu tƣơng của nƣớc
ta phát triển chậm [43].
Bảng 1.1. Tình hình sản xuất đậu tƣơng ở Việt Nam từ năm 2003 đến 2010 [42]
Năm
Chỉ tiêu
2003
2004
2005
2006
2007
2008
2009
2010
Diện tch (1000 ha)
165,6
183,8
204,1
185,6
187,4
192,1
146,2
197,8
Năng suất (tạ/ha)
13,3
13,3
14,3
13,9
14,7
14,0
14,6
15,0
Sản lƣợng (1000 tấn)
219,7
245,9
292,7
258,1
275,2
267,6
213,6
296,9
(Nguồn: Tổng cục thống kê, 2011)
V diện tch , đậu tƣơng đƣợ c gieo trng trên diệ n tí ch chỉ chiếm 1 tỉ lệ rất thấp
khoảng 1,5-1,6 %, xét v tốc độ thì diện tch gieo trng vn tăng qua các năm. Năm 1980,
diện tch trng đậu tƣơng là 48,8 nghìn ha, nhƣ vậy tnh đến năm 2010, diện tch đã tăng
7
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
lên gấp 4,05 lần. V năng suất: năng suất bình quân của đậu tƣơng ở nƣớc ta là rất thấp,
chỉ bng 50% - 70% năng suất đậu tƣơng của thế giới [42].
Vớ i sả n lƣợ ng đậ u tƣơng chƣa đủ phụ c vụ nhu cầ u , hàng năm Việt Nam vn phải
nhậ p khẩ u mộ t lƣợ ng lớ n đậ u tƣơng tƣ̀ nƣớ c ngoà i . Lƣợ ng đậ u tƣơng nhậ p khẩ u nà y , ¾
dành cho sản xuất thức ăn chăn nuôi, ¼ cò n lạ i sả n xuấ t sả n phẩ m phụ c vụ con ngƣờ i [42].
Nhƣ vậy, việ c phá t triể n diệ n tích và sả n lƣợ ng đậ u tƣơng là rấ t cầ n thiế t và yêu
cầu đt ra là phải chn tạo ra giống đậu tƣơng có năng suất cao hơn thông qua các biện
pháp truyn thống và đc biệt là việc áp dng công nghệ gen.
1.2. HẠN V TC ĐỘNG CỦA HẠN ĐẾN CÂY ĐẬ U TƢƠNG
Hạn đối với thực vật là khái niệm dùng để chỉ sự thiếu nƣớc do môi trƣờng gây
nên làm ảnh hƣởng đến sinh trƣởng và phát triển củ a cây . Những cây có khả năng duy trì
sự phát triển và cho năng suất ổn định trong điu kiện hạn gi là cây chịu hạn. Khả năng
của thực vật có thể làm giảm mức độ tổn thƣơng do thiếu nƣớc gây ra gi là tnh chịu hạn.
Có hai loại hạn cơ bản: hạn thật (gm hạn đất, hạn không kh, hạn toàn diện), và
hạn sinh l. Hạn đất là do lƣợng nƣớc trong đất giảm làm hệ rễ của cây không thể lấy
nƣớc từ đất vào tế bào, dn đến cây có hiện tƣợng bị héo lâu dài. Hạn không kh thƣờng
có đc trƣng là nhiệt độ cao (39 - 42
0
C) và độ m thấp hơn 62%. Hạn không kh thƣờng
gây nên hiện tƣợng héo tạm thời, vì rễ cây không hút đủ nƣớc mà quá trình thoát hơi nƣớc
quá nhanh do nhiệt độ môi trƣờng cao. Hạn sinh l khác với hai loại hạn trên ở chỗ môi
trƣờng vn đầy đủ nƣớc nhƣng do mất cân bng áp suất thm thấu giữa môi trƣờng tế bào
và môi trƣờng bên ngoài nên rễ cây không có khả năng hút nƣớc [10].
Hạn hán là một trong những áp lực môi trƣờng nghiêm trng và ảnh hƣởng đến
hầu nhƣ tất cả các chức năng thực vật. Cơ chế chống chịu hạn ở thực vật rất phức tạp liên
quan đến các đc điểm hình thái giải phu, sinh lí, hoá sinh, và liên quan đến các gen chịu
hạn. Hạn thƣờng làm giảm 40% năng suấ t đậ u tƣơng và tá c độ ng đế n tấ t cả cá c giai đoạ n
sinh trƣở ng và phá t triể n , tƣ̀ nả y mầ m đế n ra hoa , tạo hạt cũng nhƣ chất lƣợng hạt đƣợc
tạo thành [23], [29].
8
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
Tác đng của hạn đến hệ r : Đậu tƣơng thƣờng xuyên chịu đựng s ự thiếu nƣớc
lớ n. Thƣ̣ c vậ t nó i chung , đậ u tƣơng nó i riêng có thể thích ƣ́ ng bằ ng cá ch phá t triể n rễ
chnh dài hơn, hệ thố ng lông rễ lớ n có thể tìm nƣớ c ở cá c lớ p đấ t phía dƣớ i ẩ m hơn và hú t
dinh dƣỡ ng nhƣ phospho. Mộ t trong cá c yế u tố chính ả nh hƣở ng đế n độ sâu củ a hệ rễ đậ u
tƣơng là tỉ lệ ké o dà i rễ . Rễ cá i là dạ ng rễ đầ u củ a đậ u tƣơng, xác định kiểu hình của rễ cái
dƣớ i điề u kiệ n không stress có thể cho phé p xá c định khả năng đâm sâu củ a rễ . Kaspar và
cs đã lấ y ra 105 kiể u hình khá c nhau củ a đậ u tƣơng tƣ̀ cá c nhó m có tí nh trƣở ng thà nh
khác nhau. Độ dài của rễ cái trong mỗi nhóm trƣởng thành biến thiên vào khoảng 1,3 cm
d
-1
. Các kết quả trên cho thấ y nhó m cá c giố ng có tỉ lệ ké o dà i rễ nhanh hơn thƣờ ng có thể
hút nƣớc trong đất ở sâu 120 cm dƣớ i bề mặ t . Dƣớ i điề u kiệ n hạ n , số lƣợ ng rễ mớ i /mộ t
đơn vị chiề u dà i rễ cá i tăng lên đá ng kể , nhƣng không tăng chiề u dà i h ay đƣờ ng kính rễ
cái. Bở i vì , sƣ̣ sinh trƣở ng rễ và sƣ̣ phân phố i nƣớ c là quyế t đị nh để duy trì chƣ́ c năng
trong cá c điề u kiệ n môi trƣờ ng khá c nhau , đặ c tính mề m dẻ o củ a rễ là yế u tố quyế t định
khả năng tìm đƣợc các ngu n nƣớ c. Stress nƣớ c hƣớ ng tớ i sƣ̣ gia tăng phân chia khố i
lƣợ ng hợ p chấ t sinh họ c đế n rễ , gia tăng tỉ lệ rễ : thân. Các cây không đƣợ c tƣớ i nƣớ c cho
thấ y sƣ̣ gia tăng kích thƣớ c rễ khi so sá nh vớ i cây đƣợ c tƣớ i nƣớ c, xuấ t hiệ n tƣơng quan
đặ c trƣng giƣ̃ a sƣ́ c chịu đƣ̣ ng hạ n vớ i cá c đặ c điể m đa dạ ng củ a rễ nhƣ khố i lƣợ ng khô ,
kch thƣớc toàn bộ hệ rễ , khố i lƣợ ng và số lƣợ ng rễ phụ [23]. Phân tch proteomic của rễ
cây đậ u tƣơng xƣ̉ lý hạ n, phát hiện 35 protein đƣợ c tạ o ra và có nhiề u thay đổ i quan trọ ng
ở vùng rễ bị stress mất nƣớc khi so sánh với điu kiện bình thƣờng . Số lƣợ ng của mộ t số
protein gia tăng , nhƣ là ferritin, trong khu vƣ̣ c ké o dà i củ a rễ bị xƣ̉ lý stress , có vai trò
quan trọ ng bả o vệ tế bà o chố ng lạ i cá c dạ ng oxy hoạ t hoá . Thông tin tƣ̀ proteomic đã cho
thấ y mố i quan hệ gầ n gũ i giƣ̃ a sinh tổ ng hợ p phenylpropanoid và sinh trƣở ng củ a rễ trong
điề u kiệ n thế nƣớ c thấ p , nhƣ sƣ̣ gia tăng lớ n củ a lƣợ ng isoflavon và cá c bƣ́ c vá ch
phenolic, bao gồ m lignin lầ n lƣợ t ở vù ng chó p rễ và vù ng cơ bả n [29].
Tác đng của hạn đến khả năng c định đạm : Nitơ liên kế t trong cây họ đậ u rấ t
nhạy cảm với đất khô . Trong điề u kiệ n hạ n , cây đậ u tƣơng không chỉ bị mấ t cân bằ ng
lƣợ ng CO
2
tch lũy và làm giảm sự phát triển diện tch lá , mà lƣợng nitơ liên kết cũng
9
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
không cò n đƣợ c giƣ̃ vƣ̃ ng . Đất khô làm giảm của hàm lƣợng nitơ cho quá trình tạo ra
protein, làm giảm năng suất cây trng và chất lƣợng hạt giống . Mộ t và i yế u tố liên quan
đến ức chế nitơ liên kết trong điu kiện hạn bao gm : hạn chế lƣợng oxy đi vào ; giảm
dòng cacbon đến nốt sần ; giảm hoạt động tổ ng hợ p đƣờ ng trong nố t sầ n ; đồ ng thờ i tăng
tch lũy urê và các amino acid tự do [23], [29]. Hoạt động của nitrogenase cho thấy giảm
70% trong 4 ngày đầu gây hạn , trong khi quang hợ p chỉ giả m 5%, điề u nà y cho thấ y
stress nƣớ c tạ o mộ t tá c độ ng đế n hoạ t độ ng củ a nitrogenase , mà không ph thuộc tỉ lệ
quang hợ p. Nó cũng cho thấy sự thiếu nƣớc tác động đến hoạt động của nốt sần thông qua
sƣ̣ gia tăng khả năng chị u đƣ̣ ng sƣ̣ khuế ch tá n oxy đế n bacteriod . Khả năng chống lại sự
khuế ch tá n oxy đƣợ c tăng cƣờ ng ; giảm hô hấp nitrogenase liên kết và hoạt động các
enzyme; tch lũy các chất nn hô hấp và lipid bị oxy hóa ; điề u tiế t ngƣợ c cá c gen chố ng
lại oxy hóa; cùng với hoạt động tậ p trung hô hấ p củ a bacteriod bị là m suy yế u trong điề u
kiệ n hạ n, và sự hƣ hại oxy hóa xảy ra trong nốt sần trƣớc khi đến tác động nào tới sinh
tổ ng hợ p đƣờ ng hay leghemoglobin . Đất khô thƣờng điu khiển sự tch lũy của urê trong
lá cây đậu tƣơng và đƣợc biết tới nhƣ một nhân tố hạn chế sự tạo nốt sần [29].
Tác đng của hạn đến hnh thái lá : Lông tơ lá thƣờ ng là mộ t đặ c điể m phổ biế n
của thực vậ t chị u hạ n , cũng nhƣ mộ t và i loạ i câ y trồ ng, trong đó có đậ u tƣơng . Thông
thƣờ ng, độ dà y lông tơ lá gia tăng hệ số phả n xạ tƣ̀ lá , dẫ n đế n giả m nhiệ t độ củ a lá trong
điề u kiệ n chiế u sá ng cao. Độ dày lông tơ lá là đc điểm thch ứng quan trng cho cây đậu
tƣơng dƣớ i điề u kiệ n hạ n . Lông tơ dà y đặ c đƣợ c tăng cƣờ ng sƣ́ c sinh trƣở ng , rễ dà y đặ c
hơn và mở rộ ng xuố ng sâu hơn . Sƣ̣ gia tăng độ dà y lông tơ lá có thể là m tăng lớ p ranh
giớ i chố ng lạ i stress lên hơn 50%, giảm nhiệt độ của lá , hạn chế thoát hơi nƣớc và tăng
cƣờ ng quá trình quang hợ p. Hơn nƣ̃ a, sƣ̣ gia tăng độ dà y lông tơ có thể là m giả m đi đá ng
kể phạ m vi ả nh hƣở ng bệ nh khả m virus củ a đậ u tƣơng [29].
Tác đng củ a hạ n đế n đặ c điể m sinh lý củ a t hân: Mộ t trong nhƣ̃ ng đặ c điể m
sinh lý liên quan đế n thân có thể tá c độ ng đế n khả năng chịu hạ n là sƣ̣ suy giả m lƣợ ng
nƣớ c sƣ̉ dụ ng trong toà n bộ cơ thể thƣ̣ c vậ t khi đấ t bị thiế u nƣớ c . Khi sƣ̣ mấ t nƣớ c ở đấ t
gia tăng, thƣ̣ c vật phải chịu đựng một thời kỳ chuyển tiếp từ trạng thái bão hòa nƣớc , khi
10
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
mà lƣợng nƣớc toàn bộ cơ thể thực vật sử dng không ph thuộc vào lƣợng nƣớc trong
đấ t; sang trạ ng thá i thƣ́ hai khi mà nƣớ c sƣ̉ dụ ng liên quan tr ực tiếp tới lƣợng nƣớc dùng
đƣợ c trong đấ t . Việ c chuyể n trạ ng thá i nà y đƣợ c kế t hợ p vớ i sƣ̣ giả m chỉ số dẫ n truyề n
trung bình ở khí khổ ng, và có thể xảy ra ph thuộc lƣợ ng nƣớ c trong đấ t khá c nhau, trong
nhiề u loà i khá c nhau. Tnh dn truyn của kh khổng tác động quan trng đến sự thay đổi
sƣ̣ trao đổ i khí củ a lá và thoá t hơi nƣớ c . Các tác động của hạn đến sinh trƣởng của lá , sƣ̣
dẫ n truyề n củ a khí khổ ng và mố i quan hệ vớ i lƣợ ng n ƣớc trong cây đã đƣợc nghiên cứu
trong cây đậ u tƣơng . Nó cho thấy rng hạn làm giảm tƣơng đối tỉ lệ mở rộng của lá , sƣ̣
dẫ n truyề n củ a khí khổ ng và sƣ́ c căng củ a lá , trong khi đó nó tăng cƣờ ng lƣợ ng abscisis
acid trong lá và mạch xylem. Sƣ̣ giả m tí nh dẫ n củ a khí khổ ng xả y ra đồ ng thờ i vớ i sƣ̣ gia
tăng abscisis acid trong xylem và xả y ra trƣớ c bấ t cƣ́ sƣ̣ thay đổ i quan trọ ng nà o về sƣ́ c
căng củ a lá , nó chỉ ra rng các dấu hiệu hóa hc (abscisis acid - nguồ n gố c tƣ̀ rễ ) điề u
khiể n hoạ t độ ng khí khổ ng ở trạ ng thá i mấ t nƣớ c vƣ̀ a phả i [9], [29].
Tác đng của hạn đến áp suất thẩm thấu : Khả năng điu chỉnh áp suất thm
thấu có mối liên quan trực tiếp đến khả năng cạnh tranh nƣớc của tế bào rễ cây đối với
đất. Trong điu kiện hạn, áp suất thm thấu tăng lên giúp cho tế bào rễ thu đƣợc các phân
tử nƣớc hiếm hoi có trong đất nhờ vậy thực vật có thể vƣợt qua tình trạng hạn cc bộ. Khi
tế bào mất nƣớc, các chất hoà tan sẽ đƣợc tch lu nhm chống lại việc giảm mất nƣớc,
tăng khả năng giữ nƣớc của nguyên sinh chất và chúng có thể thay vị tr nƣớc nơi xảy ra
các phản ứng sinh hoá, tƣơng tác với lipid hoc protein trong màng ngăn chn sự phá huỷ
của màng và các phức protein. Các chất hoà tan có khả năng tạo áp suất thm thấu cao
bao gm các ion K
+
, proline, acid hữu cơ , các loại đƣờng . Khi bắt đầu mất nƣớc , hàm
lƣợng proline và abscisic acid tăng , tăng tổng hợp protein, nhƣng nếu hạn kéo dài sẽ xảy
ra hiện tƣợng thuỷ phân protein. Quá trình thuỷ phân các hydratcacbon dự trữ cũng là
ngun cung cấp chất tan cho quá trình điu chỉnh áp suất thm thấu [23], [29].
Abscisic acid (ABA) đƣợc tạo ra dƣới stress mấ t nƣớc và đóng vai trò quan trng
trong việc chống chịu đối với hạn hán. Trong trƣờng hợp có mt ABA quá trình đóng mở
lỗ kh đƣợc điu chỉnh làm hạn chế sự thoát hơi nƣớc. Hầu hết gen cảm ứng stress hạn
11
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
cũng đƣợc gây ra bởi tác động ABA (Shinozaki và Yamaguchi-Shinozaki năm 1997,
2000). Tuy nhiên, trong các thể đột biến Arabidopsis aba (thiếu ABA) hoc abi (không
nhạy cảm với ABA) có một số gen khác đã đƣợc gây ra bởi hạn hán, muối và lạnh. Điu
này cho thấy một số gen lại không yêu cầu ABA cho biểu hiện của chúng dƣới các điu
kiện mn, hạn hán và lạnh [25].
1.3. CC NHÓM GEN LIÊN QUAN ĐẾ N TÍ NH CHỊ U HẠ N
1.3.1. Các nhm gen liên quan đến tính chịu hạn
Hạn, mặ n và lạ nh là m giả m tình trạ ng nƣớ c trong tế bà o thƣ̣ c vậ t và gây tổ n
thƣơng cho cây, có thể dn đến chết cây . Ở mức độ phân tử , điề u kiệ n hạ n sẽ là m cho
thƣ̣ c vậ t gia tăng mƣ́ c độ biể u hiệ n và tích lũ y củ a cá c gen /protein chố ng chịu stress. Một
số gen đƣợc tăng hoạt động và một số khác bị giảm hoạt động trong cây bị stress hạn. Sự
điu chỉnh này xảy ra ở các mức riêng biệt, từ thời điểm phát hiện stress đến việc sản xuất
các protein hoạt tnh sinh hc (Shinozaki và Yamaguchi-Shinozaki, 2007). Các cơ chế
bảo vệ tham gia vào quá trình này là khá giống nhau trong giới thực vật. Do đó, nghiên
cứu sử dng cây mô hình, nhƣ A.thaliana và thuốc lá (Nicotiana tabacum), có thể giúp
xác định những gen có chức năng quan trng trong cơ chế bảo vệ trong các loài thực vật
khác. Các gen mã hóa protein tham gia và o phả n ƣ́ ng củ a thƣ̣ c vậ t vớ i cá c tá c nhân stress đƣợ c
phân thà nh hai nhóm dựa vào chức năng của các sản phm mà chúng mã hóa [8], [19], [25].
(1) Nhóm protein chức năng tham gia trực tiếp đáp ứng điu kiện hạn , bảo vệ các
đạ i phân tƣ̉ và mà ng (protein LEA, osmotin, protein chố ng đông , chaperonin và protein
liên kế t mRNA…) và duy trì nƣớc tự do qua màng (protein kênh vậ n chuyể n nƣớ c và cá c
kênh xuyên mà ng…), các enzyme xúc tác các quá trình sinh tổng hợp các chất thm thấu
(proline, betain và đƣờ ng ) và các enzyme khƣ̉ độ c cho phé p cá c hoạ t độ ng sinh lý trong
tế bà o hay sƣ̣ chuyể n hó a sinh họ c duy trì mƣ́ c bình thƣờ ng (glutathinone S-transferase,
epoxide hydrolase hò a tan, catalase, superoxide và ascorbic peroxidase…) [16], [25]. Khả
năng chố ng chịu vớ i hạ n hay mặ n có thể đƣợ c tăng cƣờ ng bằ ng cá ch chuyể n gen mã hó a
protein LEA, protein trong tổ ng hợ p hoặ c tổ ng hợ p betain…[25].
12
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
(2) Nhóm protein điu khiển sự biểu hiện của các gen chức năng liên quan đến tnh
chịu hạn , các nhân tố phiên mã bá m và o trình tƣ̣ DNA đặ c hiệ u trên vù ng khở i độ ng
(promoter) của các gen chức năng tham gia vào tnh chịu hạn và hoạt hóa sự biểu hiện của
các gen này và kết quả là tăng cƣờng tnh chịu hạn của thực v ật. Các nghiên cứu v gen
đã chƣ́ ng minh vù ng khở i độ ng củ a nhiề u gen chƣ́ c năng chƣ́ a mộ t hoặ c nhiề u trình tƣ̣
DNA đặ c hiệ u là điể m bá m củ a mộ t nhân tố phiên mã . Điề u nà y chƣ́ ng tỏ mộ t nhân tố
phiên mã có thể hoạ t hó a biể u hiệ n củ a nhiề u gen chƣ́ c năng . Các nghiên cứu biểu hiện
gen đã chƣ́ ng minh nhiề u nhân tố phiên mã biể u hiệ n mạ nh trong điề u kiệ n hạ n , chƣ́ ng tỏ
nhóm protein này đóng vai trò quan trng trong cơ chế điu hòa và biểu hiện của các gen
tăng cƣờ ng tính khá ng hạ n ở thƣ̣ c vậ t [8]. Sản phm gen thuộc nhóm hai bao gm các yếu
tố phiên mã (bZIP, MYC, MYB và DREB… ); protein kinase (MAP kinase và CDP
kinase, protein kinase thụ quan, protein kinase ribosome và protein kinase liên quan phiên
mã…); proteinase (photphoeterase và phospholipase C…) [25], [30].
1.3.2. Các nhân t phiên m điều khiển các gen liên quan đến tính chịu hạn
Quá trình từ DNA đến protein trải qua quá trình rất quan trng là phiên mã. Các
nhân tố phiên mã bá m và trình tƣ̣ DNA đặ c hiệ u trên vù ng khở i độ ng củ a cá c gen đích và
điề u hò a biể u hiệ n củ a cá c gen nà y.
Nhóm gen mã hóa các nhân tố phiên mã chiếm khoảng 8-10% trong hệ gen mỗ i
loài và đóng vai trò quan trọ ng trong mọ i hoạ t độ ng số ng nhƣ quá trình sinh trƣở ng phá t
triể n và chố ng chịu vớ i bấ t lợ i môi trƣờ ng. Ở đậ u tƣơng, hơn 2000 yế u tố phiên mã tƣ̀ cá c
mô và cơ quan chính đã đƣợ c xá c định khoả ng trên 180 yế u tố phiên mã ở rễ liên quan
tnh chịu hạn [29]; hệ gen củ a Arabidopsis có khoảng 5,9% mã hóa cho trên 1500 yế u tố
phiên mã [10], [34].
Các yếu tố phiên mã liên quan đến phản ứng lại stress của thực vật đƣợc chia thành
các nhóm nhƣ là: AP2/ERF, NAC, MYB, MYC, Cys2His2 và WRKY [31]. Đc điểm đc
trƣng củ a cá c protein điề u khiể n (nhân tố phiên mã ) là có hai vùng hoạt động : vùng hoạt
hóa các protein chức năng và vùng gắn với các trật tự DNA đc hiệu trên promoter củ a
gen [9]. Các nhân tố phiên mã điu khiển các gen chịu hạn phân loại dựa vào trật tự DNA
13
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
đặ c hiệ u mà chú ng liên kế t. Vùng khởi động (promoter) của hầu hết các gen liên quan đến
tnh chịu hạn thƣờng chứa mộ t hoặ c nhiề u trậ t tƣ̣ DNA đặ c hiệ u phổ biế n là ABRE (yế u
tố đá p ƣ́ ng ABA ) và DRE/CRT (yế u tố đá p ƣ́ ng hạ n /đoạ n lặ p C ), NAC, MYB… vớ i
chƣ́ c năng tƣơng ƣ́ ng là điể m bá m củ a cá c protein điề u khiể n quá trình phiên mã biể u hiệ n
ph thuộc và không ph thuộc vào ABA [8], [19].
Nhm nhân t phiên m không ph thuc ABA
NAC: các nhân tố phiên mã h NAC chứa trình tự đng nhất gi là vùng hoạt động
NAC ở đầ u N . Đây là mộ t họ gen đặ c trƣng ở thƣ̣ c vậ t, có vai trò quan trng trong việc
xác định mô phân sinh đỉnh chi, biệ t hó a cá c cơ quan rễ , hoa trong sinh trƣở ng phá t triể n
thƣ̣ c vậ t, phản ứng lại điu kiện bị tổn thƣơng và tác nhân gây hại tấn công [9]. Hệ gen
Arabidopsis có t nhất 100 gen họ NAC, và hệ gen lúa có 75 gen đã đƣợ c phân lậ p nhƣng
chỉ một số t gen trong số này đƣợc nghiên cứu chức năng [34], ở đậu tƣơng cũng có hơn
100 gen NAC đƣợ c tìm thấ y trong hệ gen [30]. Phầ n lớ n protein củ a cá c gen trong họ
NAC có chƣ́ a mộ t vù ng liên kế t DNA ở tậ n cù ng đầ u N có độ bả o thủ cao, mộ t trình tƣ̣ tín
hiệ u định vị nhân và mộ t vù ng tậ n cù ng đầ u C biế n đổ i.
Nhm nhân t DREB: trình tự đc hiệu của nhóm nhân tố DREB1/CRT có trình tự
li là A /GCCGAC lầ n đầ u tiên đƣợ c phá t hiệ n nằ m trong vù ng điề u khiể n gen rd29 ở
Arabidopsis và sau này đƣợc phát hiện trên rất nhiu vùng điu khiển gen của các gen
biể u hiệ n trong điề u kiệ n hạ n và l ạnh khác. Các gen điu khiển quá trình phiên mã thuộc
nhóm AP2 (APETALA2)/ethylene – responsive element-binding factor (ERF) bám vào
trình tự DRE và đƣợc đt tên là : DREB1/CBF và DREB2 [19]. AP2/ERF là mộ t họ lớ n
bao gồ m cá c yế u t ố phiên mã quan trng . Các protein AP 2 của Arabidopsis đƣợ c chia
thành 5 nhóm dựa vào sự giống nhau trong vùng gắn DNA của chúng : h AP2 (14 gen),
h RAV (liên quan tớ i ABI3/VP1) (6 gen), h DREB (55 gen; nhóm A), h ERF (65 gen,
nhóm B), và các gen khác (4 gen), mộ t gen rấ t đặ c biệ t AL079349 [14], [15], [16], [18]. Ở
đậ u tƣơng cũ ng có 5 nhóm gen tƣơng tự , hiệ n nay có hơn 380 gen AP2/ERF đƣợ c phá t
hiệ n ở hệ gen đậ u tƣơng [30]. Các protein h AP 2 chƣ́ a đƣ̣ ng hai vùng AP2/ERF, và các
gen trong họ nà y tham gia điề u tiế t cá c quá trình phá t triể n . H protein RAV chứa một
14
Số hóa bởi Trung tâm Học liệu – Đại học Thái Nguyên
vùng AP2/ERF và mộ t vù ng B3, cái mà khác với các chức năng sinh hc và liên quan đến
các loại khác của quá trình ph iên mã . Trong thƣ̣ c tế , các thành viên h RAV liên quan
phản ứng lại ethylene , brassinosteroid, và các stress môi trƣờng và sinh hc . Đối lập với
các thành viên h AP 2 và RAV, các protein h DREB/CBF và ERF chỉ chƣ́ a mộ t vù ng
AP2/ERF. Hệ gen Arabidopsis mã hóa 145 protein liên quan DREB/CBF. Các gen của h
DREB/CBF giƣ̃ vai trò quyế t định trong sƣ̣ chố ng lạ i stress vô sinh ở thƣ̣ c vậ t bằ ng cá ch
nhậ n ra nhân tố phả n ƣ́ ng lạ i sƣ̣ mấ t nƣớ c và lạ nh (DRE/CRT) vớ i mô hì nh trung tâm
A/GCCGAC. H ERF liên quan chnh đến phản ứng lại stress sinh hc nhƣ khi phát sinh
bệ nh bằ ng cá ch nhậ n ra nhân tố hoạ t độ ng cis AGCCGCC hay gọ i là hộ p GCC . Nhiề u
thành viên h ERF cũng gắn với các nhân tố DRE/CRT [39].
Hnh 1.1. Hệ thống nhân tố phiên mã điề u khiể n sƣ̣ biể u hiệ n cá c gen chƣ́ c năng tham
gia phả n ƣ́ ng chố ng chịu vớ i điề u kiệ n bấ t lợ i củ a Arabidopsis [8]
Các nhân t phiên m ph thuc ABA
Nhm nhân t ABRE /ABF: trậ t tƣ̣ DNA đặ c hiệ u ABRE có trình tƣ̣ lõ i
ACGTGGC, lầ n đầ u tiên đƣợ c phá t hiệ n nằ m trên vù ng điề u khiể n gen Em ở lúa mỳ, gen
rab16 ở lúa. Hai nhó m protein điề u khiể n quá trình phiên mã ABRE /ABF bá m và o trậ t tƣ̣