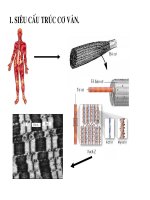
BÀI GIẢI SINH HÓA (P2)
Bạn đang xem bản rút gọn của tài liệu. Xem và tải ngay bản đầy đủ của tài liệu tại đây (580.95 KB, 20 trang )
Chương 5: CARBOHYDRATE
1. Trong các phân tử sugar alcohol (các dẫn xuất của monosaccharide), nguyên tử
oxy ở nhóm carbonyl bị khử thành nhóm hydroxyl, ví dụ D-glyceraldehyde có thể
bị khử thành glycerol. Tuy nhiên, hợp chất sugar alcohol này không có dạng đồng
phân D hoặc L. Giải thích?
Khi nhóm carbonyl bị khử thành hydroxy (-OH)l, C thứ 2 (gắn với nhóm carbonyl)
không còn là Carbon bất đối xứng nữa vì có 2 nhóm -CH2OH. Vì thế, phân tử này
không có đồng phân D/L.
Glyceraldehyde
2. Nhiều carbohydrate phản ứng với phenylhydrazine (C
6
H
5
NHNH
2
) tạo thành
những dẫn xuất có tinh thể màu vàng tươi, được gọi là osazone:
Nhiệt độ nóng chảy (melting point-MP) của những dẫn xuất này được xác định rất
dễ dàng và phụ thuộc đặc tính từng osazone. Thông tin này được dùng để xác định
các monosaccharide trước khi cho qua HPLC hoặc sác ký khí-lỏng. Bảng dưới liệt
MP của một vài dẫn xuất aldoseosazone:
Monosaccharid
e
MP của
monosaccharide khan
(
o
C)
MP của dẫn xuất
osazone (
o
C)
Glucose 146 205
Mannose 132 205
11
Galactose 165-168 201
Talose 128-130 201
Như đã cho trên bảng, từng cặp
dẫn xuất osazon của glucose và
mannose, cùng với galactose và
talose, có cùng điểm nóng chảy
theo từng cặp mặc dù các
monosaccharide đơn của chúng
thì có điểm nóng chảy khác nhau.
Giải thích?
Phản ứng giữa phân tử đường và phenylhydrazine xảy ra tại vị trí nhóm carbonyl
C1 và hydroxyl C2 của phân tử đường. Các cặp đường glucose và mannose,
galactose và talose là các đồng phân lập thể tương ứng của nhau (epimer) chỉ
khác nhau về cấu hình tại vị trí C2 của từng cặp đường này, vì vậy phản ứng
osazone tạo ra các dẫn xuất giống nhau h oàn toàn về công thức cấu tạo cũng như
cấu hình 3D. Vì vậy nhiệt độ nóng chảy của các cặp dẫn xuất osazone giống nhau.
3. Dung dịch một chất đồng phân lập thể của một monosaccharide cho trước quay
mặt phẳng phân cực ánh sáng sang trái (ngược chiều kim đồng hồ) được gọi là
đồng phân dạng L (levo-rotatory – L) ký hiệu (-); những đồng phân lập thể khác
cũng quay mặt phẳng phân cực ánh sáng với cùng phạm vi như trên nhưng sang
phải (cùng chiều kim đồng hồ) được gọi là đồng phân dạng D (dextro-rotatory –
D) ký hiệu (+). Một hỗn hợp với số mol bằng nhau của hai dạng (+) và (-) sẽ
không làm quay mặt phẳng phân cực ánh sáng.
Hoạt tính quang học của các đồng phân lập thể được biểu thị một cách định lượng
thông qua tính quay cực quang học của nó, góc mà mặt phẳng phân cực ánh sáng
quay khi dung dịch một chất được đặt vào ánh sáng có bước sóng nhất định sẽ cho
biết nồng độ của chất đó. Góc quay riêng
[ ]
C
D
o
25
α
của một hợp chất có hoạt tính
quang học được xác định như sau:
22
[ ]
( )
( ) ( )
mLgCdmOPL
OOR
o
C
D
o
/
25
×
=
α
OOR (observed optical rotation): góc quay quang học quan sát được
OPL (optical path length): chiều dài sóng ánh sáng truyền qua dung dịch
C (concentration): nồng độ dung dịch
Nhiệt độ và bước sóng của ánh sáng sử dụng (thông dụng là ánh sáng Natri, bước
sóng 589nm) phải được xác định rõ trong định nghĩa.
Một dung dịch mới pha của α-D-glucose có góc quay riêng là +112
o
C. Sau một
thời gian, góc quay này giảm dần và đạt giá trị cân bằng tương ứng với
[ ]
o
C
D
o
5.52
25
+=
α
. Ngược lại, dung dịch mới pha của β-D-glucose có góc quay cực
riêng là +19
o
. Sự quay cực của dung dịch này theo thời gian tăng dần đến giá trị
cân bằng như ở đồng phân α.
a- Vẽ công thức phối cảnh Haworth của hai dạng đồng phân α và β của D-
glucose. Điểm khác nhau đặc biệt của hai dạng đồng phân trên là gì?
Cấu trúc của phân tử đường vẽ trên mặt phẳng là dạng cấu trúc Fisher
Cấu trúc vẽ trên mặt phẳng của phân tử đường dạng vòng là dạng cấu trúc
Haworth
cấu
trú
c
Fis
her
cấu
trú
c
Ha
wo
rth
α-
D-
glu
cấu
trú
c
Ha
wo
rth
β-
D-
glu
33
của
D-
glu
cos
e
cop
yra
nos
e
cop
yra
nos
e
Ở đây đặc trưng
α
và
β
là cấu hình quay của nhóm hydroxyl (-OH) quang C- khử
(hay gọi) là hemicetal của vòng pyranose
b- Tại sao góc quay riêng của dung dịch mới pha ở dạng đồng phân α lại
giảm theo thời gian? Tại sao dung dịch hai dạng đồng phân α và β lại có cùng giá
trị góc quay riêng ở trạng thái cân bằng?
Trong dung dịch D-glucose, quá trình chuyển đổi đồng phân qua lại
(mutarotation) luôn dẫn đến luôn có sự tồn tại 2 dạng đồng phân quang học α và
β. Hỗn hợp đồng phân này trở nên cân bằng về đặc tính quang học trong dung
dịch. Điều đó có nghĩa rằng dung dịch ban đầu chỉ toàn phân tử α hoặc β, nhưng
sau một thời gian 2 kiểu dung dịch này trở thành hỗn hợp cân bằng, tương đồng
nhau về góc quay quang học.
Dung dịch ban đầu chỉ toàn α-glucose
[ ]
C
D
o
25
α
= +112
o
thì sau một thời gian
một số phân tử chuyển sang dạng β-glucose
[ ]
C
D
o
25
α
= +19
o
gây nên việc
giảm góc quay chung của dung dịch.
c- Tính toán phần trăm mỗi loại đồng phân trên có mặt trong dung dịch D-
glucose ở trạng thái cân bằng
Giả sử có 100 phân tử D-glucose trong dung dịch, mỗi phân tử α-glucose có góc
quay cực riêng là +112
o
, mỗi phân tử β-D-glucose có góc quay cực riêng là +19
o
.
Góc quay cực của dung dịch (bao gồm phân tử α-glucose và β-D-glucose) là
44
trung bình cộng của các góc quay cực riêng của các phân tử trong dung dịch. Do
đó tại trạng thái cân bằng ta có hệ phương trình 2 nghiệm thức:
X
1
+ X
2
= 100 (%)
112.X
1
+ 19.X
2
= 52.5(X
1
+ X
2
)
Từ phương trình ta tính được X
1
=36; X
2
=64 hay nói cách khác tại trạng thái cân
bằng α-D-glucose chiếm 36% và β-D-glucose chiếm 64%
4. Fructose trong mật ong tồn tại chính ở dạng β-D-pyranose. Đây là một trong
những carbohydrate ngọt nhất được biết tới, có độ ngọt khoảng gấp đôi độ ngọt của
glucose. Dạng β-D-furanose của fructose thì lại ít ngọt hơn. Độ ngọt của mật ong
giảm ở nhiệt độ cao. Cũng như vậy, nước mật bắp giàu fructose (một sản phẩm
thương mại trong đó glucose của dịch mật bắp được chuyển sang dạng fructose)
được dùng trong những thức uống lạnh chứ không dùng cho nước uống nóng. Đặc
tính hóa học nào của fructose có thể giải thích cho những điều trên?
Fructose có khả năng đóng vòng tạo 2 dạng β-D-pyranose và β-D-furanose. Khi
nhiệt độ cao, cân bằng xảy ra theo hướng tạo nhiều phân tử β-D-furanose hơn,
dẫn đến giảm độ ngọt của dung dịch fructose.
5. Thủy phân sucrose (góc quay riêng là +66.5
o
) thu được hỗn hợp D-glucose (góc
quay cực riêng là +52.5
o
) và D-fructose (góc quay cực riêng là –92
o
) với số mol
bằng nhau.
a- Hãy đề xuất quy trình hợp lý để xác định mức độ thủy phân sucrose bởi
một dịch enzyme được trích từ niêm mạc ruột non.
Hoạt tính enzyme invertase (thủy phân sucrose) thể hiện bằng lượng sản phẩm tạo
thành khi enzyme thủy phân cơ chất sucrose trong một đơn vị thời gian. Như vậy,
nếu tính được lượng glucose hay fructose tạo thành trong thời gian nhất định, có
thể suy ra được hoạt tính enzyme.
Quy trình: cho enzyme tác dụng với dung dịch sucrose và đo sự thay đổi góc quay
riêng theo thời gian.
b- Giải thích tại sao hỗn hợp D-glucose và D-fructose với số mol bằng nhau
trên được gọi là đường nghịch đảo (invert sugar) trong công nghiệp thực phẩm?
55
Dung dịch sucrose ban đầu có góc quay cực dương (+66.5
o
), sau khi thuỷ phân
tạo hỗn hợp glucose và fructose với số mol bằng nhau, có góc quay cực trung bình
là: +52.5 + (-92)=-39.5
o
. Như vậy sau khi thủy phân dung dịch bị đảo cực góc
quay, và hỗn hợp dung dịch lúc này gọi là đường nghịch đảo.
c- Invertase (tên thông dụng là sucrase) được cho phản ứng với dung dịch
10% (0.1g/mL) sucrose cho tới khi sự thủy phân xảy ra hoàn toàn. Góc quay cực
quang học quan sát được trong dung dịch có bề dày 10cm? (Bỏ qua tác động nhỏ
có thể có từ enzyme).
Áp dụng công thức tính góc quay quang học:
OOR (observed optical rotation): góc quay quang học quan sát được
OPL (optical path length): chiều dài đường ánh sáng truyền qua dung dịch
(cuvette) = 10cm = 1dm
C (concentration): nồng độ dung dịch
Vì phản ứng thủy phân hoàn tòan sucrose, nên sau phản ứng trong dung dịch có
chứa cả 2 loại phân tử D-glucose và D-fructose với số lượng phân tử ngang nhau
(tỷ lệ 1:1). Do đó góc quan sát được sau phản ứng sẽ là trung bình cộng góc quay
quan sát được của dung dịch
6. Mặc dù lactose tồn tại dưới hai dạng đồng phân anomer, nhưng không tìm thấy
dạng anomer nào của sucrose. Giải thích?
Anomer là đồng phân cấu hình tại C có chứa nhóm hemiacetal hay hemiketal.
Lactose cấu tạo từ 2 phân tử β-galactose và β-glucose liên kết bằng cầu nối 1,4 vì
thế lactose còn dư nhóm hemiacetal trong phân tử
có 2 đồng phân
66
Sucrose cấu tạo từ α-glucose và β-fructose theo cầu nối 1,2 vì thế sucrose không
còn nhóm hemiacetal tự do trong phân tử
không có đồng phân anomer
7. Hầu hết cellulose tinh khiết thu được từ hạt Gossypium (cotton) mang các đặc
tính dai, dạng sợi, và không hòa tan trong nước. Ngược lại, glycogen thu được từ
cơ hoặc gan lại phân tán dễ dàng trong nước nóng tạo dung dịch đục. Mặc dù có
những tính chất vật lý khác nhau nhưng chúng đều là polymer được cấu tạo từ liên
kết (1→4) D-glucose với trọng lượng phân tử có thể ngang bằng với nhau. Những
khác biệt nào về cấu trúc phân tử làm cho hai polysaccharide trên có tính chất vật
lý khác nhau? Giải thích những thuận lợi về mặt sinh học từ tính chất những hợp
chất trên.
Cellulose cấu tạo từ các đơn phân β-glucose nối với nhau theo cầu nối β-1,4
glycoside. Cầu nối này làm cho 2 phân tử glucose kế cận nhau xoay 180
o
, liên kết
này cứng nhắc nên 2 phân đường khó có thể quay xung quanh trục liên kết. Các
đơn phân β-glucose khác tiếp tục gắn song song vào mạch tạo nên cấu trúc
polymer thảng dài, dày, chặt chẽ. Chính cấu trúc cứng nhắc này làm cho cellulose
không tan được trong nước.
Glycogen cấu tạo từ các đơn phân α-glucose nối với nhau theo cầu nối α-1,4
glucoside. Cầu nối này làm phân tử glucose thứ 2 quay một góc 60
o
so với phân tử
glucose trước đó. Giữa 2 phân từ đường vẫn có thể quay xung quanh trục liên kết
dẫn đến cấu trúc polymer có khuynh hướng sợi gấp khúc, gấp cuộn. Vì vậy,
glycogen có mạch không dài, dạng hạt, có khả năng phân tán trong nước.
Thuận lợi về mặt sinh học: cellulose do cấu trúc cứng nhắc và thẳng nên có vai trò
thành tế bào thực vật (mô xơ, sợi, mạch gỗ). Glycogen dạng hạt, gọn về thể tích,
nên thích hợp tích lũy trong mô của động vật
8. Thân tre (một loại cỏ nhiệt đới) có thể phát triển với tốc độ phi thường
0.3m/ngày trong điều kiện tối ưu. Cho rằng thân tre được cấu tạo hoàn toàn từ sợi
cellulose theo chiều phát triển của cây, hãy tính số lượng đơn phân đường cần phải
Cellulose
77
gắn vào theo chiều phát triển của sợi cellulose bằng enzyme trong một giây để đạt
được tốc độ phát triển trên. Mỗi phân tử D-glucose chiếm khoảng 0.5nm chiều dài
của phân tử cellulose.
Tốc độ phát triển trong 1 giây: 0,3m/ngày = 0,3.10
9
nm/(24*3600)s = 3472 nm/s
Số đơn phân D-glucose cần gắn trong 1 giây: 3472/0,5=6944 (nhóm)
Bài tập chương 5 (tiếp): Chu trình đường phân (glucolysis), TCA, pentophosphate và
tổng hợp đường (gluconeogenesis)
1. Định nghĩa quá trình “Glycolysis”, quá trình này xảy ra ở điều kiện hiếu khí hay kị
khí?
Quá trình Glycolysis là quá trình phân giải phân tử glucose 6C thành hai phân tử
pyruvate 3C bằng các enzyme thông qua 1 loạt các phản ứng xúc tác bởi enzyme,
trong điều kiện kỵ khí
2. Trong quá trình chuyển hóa glucose kị khí, 1 mole Glc chuyển thành 2 mole lactate có
kèm theo sự tạo thành phân tử gì?
Quá trình chuyển hóa glucose thành lactate thông qua quá trình glycolysis tạo
thành pyruvate, và từ pyruvate mới tạo thành lactate. Do vậy quá trình này tạo ra
2 ATP (chu trình glycolysis -2ATP + 4ATP). Mặc dù 2 NADH tạo thành từ
glycolysis nhưng đã sử dụng để chuyển hóa 2 pyruvate thành 2 lactate
Người ta gọi ATP sinh ra từ quá trình glycolysis là phosphoryl hóa mức độ cơ
chất- khác với Phosphoryl hóa oxy hóa, tức là sinh ATP do sự nhường điện tử cho
oxy
PTPƯ:
88
3. Viết phản ứng oxi hóa khử của NADH và chỉ rõ đâu là dạng khử đâu là dạng oxy hóa?
NAD
+
+ 2e + 2H
+
NADH + H
+
NADH + H
+
+ ½ O NAD
+
+ H
2
O
NAD
+
nhận điện tử : dạng oxy hóa
NADH nhường điện tử: dạng khử
4. Trong quá trình lên men của nấm men tạo etanol, nếu môi trường lên men không có
ion phosphate thì không có ethanol tạo thành. Khi cho một chút Pi, lập tức ethanol bắt
đầu được tạo thành, giải thích?
Lên men là quá trình phân giải kỵ khí hợp chất glucose tạo năng lượng, trong đó có
sản sinh hợp chất ethanol. Quá trình này bao gồm cả quá trình glycosis tạo tiền chất
cho quá trình lên men ethanol là pyruvate. Quá trình tạo thành pyruvate từ phân tử
glucose trài qua giai đoạn chuẩn bị là phosphoryl hóa glucose, vì thế cần có ion
Phosphate thì quá trình glycolysis mới xảy ra và đồng nghĩa với khả năng lên men tạo
ethanol xảy ra.
5. Những enzym nào đóng vai trò trong cả 2 quá trình: Glucolysis (phân rã Glc) và
Gluconeogenesis (sinh tổng hợp Glc)?
Các enzyme cần chung cho cả 2 quá trình glycolysis và gluconeogenesis:
glyceraldehydes 3-phosphate dehydrogenase, phosphoglycerate kinase,
99
phosphoglycerate mutase, enolase, aldolase (fructose 1,6-P aldolase), phosphohexose
isomerase
6. Enzym nào đóng vai trò trong chuyển hóa glycogen thành đường đơn glucose?
Enzyme glycogen phosphorylase.
7. Một hậu quả của việc đói là sự giảm khối lượng các mô. Vậy chuyện gì đã xảy ra với
protein của các mô cơ đó?
Khi các mô lâm vào tình trạng đói, quá trình gluconeogenesis (hình thành phân tử đường
mới) để tổng hợp glucose từ các tiền chất không phải là carbonhydrate sẽ xảy ra. Protein
trong mô sẽ bị phân giải tạo các amino acid- tiền chất cho quá trình gluconeogenesis.
8. Một quy trình phổ biến để đánh giá xem một chất có phải là tiền chất của glucose
trong động vật hữu nhũ không là bỏ đói nó cho đến khi glycogen trong gan được sử
dụng hết, sau đó đưa vào cơ thể sinh vật hợp chất cần nghiên cứu. Chất này nếu là tiền
chất của glucose thì sẽ làm tăng lượng glycogen trong gan (được gọi là glucogenic) vì
chúng phải qua bước biến đổi thành glucose 6-phosphate. Hãy đánh giá những chất
cho dưới đây có phải là glucogenic?
a- Succinate,
-
OOC-CH
2
-CH
2
-COO
-
b- Glycerol, HO-CH
2
-CH(OH)-CH
2
-OH
c- Acetyl CoA, CH
3
-CO-S-CoA
d- Pyruvate, CH
3
-CO-COO
-
e- Butyrate, CH
3
-CH
2
-CH
2
-COO
-
1010
a- succinate – hợp chất 4 carbon-muốn thành tiền chất của Glc thì cần qua chu trình
TCA để thành oxaloacetate. Tiếp đến, oxaloacetate được chuỵển hóa thành
phosphoenolpyruvate (PEP) nhờ enzym PEP carboxykinase có trong ty thể hoặc ở
cytosol (xem slide chuyển hóa pyruvate, chương 5). PEP là một chất chuyển hóa trung
gian chính của chu trình gluconeogenesis (xem chương 5).
b- glycerol –hợp chất 3 carbon-(xem chương 7 phần chuyển hóa chất béo) chuyển hóa
qua 3 bước thành glyceraldehyde-3-phosphate là một chất chuyển hoá trung gian quan
trọng của glycolysis và gluconeogenesis (xem chương 5)
c- Acetyl-CoA mặc dù là sản phẩm chuyển hóa của pyruvate bởi enzym pyruvate
hydrogenase nhưng phản ứng này không thuận nghịch, nên Acetyl-coA không chuyển
hóa thành chuyển thành pyruvate. Tuy nhiên theo chu trình TCA nếu Acetyl –CoA có thể
chuyển hóa thành oxaloaceate (xem chương 6) rồi thành PEP (xem chương 5) nên cũng
có thể là tiền chất của quá trình tổng hợp glucose
d- pyruvate là tiền chất tổng hợp Glc vì nó là hợp chất trung gian quan trọng của cả 2
quá trình glycolysis và gluconeogenesis (xem chương 5)
e- butyrate không phải là tiền chất của quá trình gluconeogenesis
9. Sự thiếu hụt bẩm sinh enzym fructose1,6-bisphosphatase trong gan làm hàm lượng
lactate trong huyết tương cao hơn mức bình thường. Giải thích.
Theo sơ đồ trên (câu 8), lactate từ mô cơ sẽ đi vào máu, để được vận chuyển đến gan.
Đến gan,lactate chuyển hóa thành pyruvate và đi vào quá trình gluconeogenesis tạo
glucose máu. Enzyme fructose 1,6-bisphosphatase có vai trò trong chuỗi phản ứng
chuỵển hóa tạo glucose từ pyruvate, vì thế nếu không có enzyme này thì lactate không
được chuyển hóa bởi gluconeogenesis thành glucose máu. Lactate không bị chuyển
hóa, ứ lại trong máu với hàm lượng cao hơn mức bình thường.
(Lưu ý: lactate tích lũy gây độc cho mô, tạo ra do quá trình khử pyruvate)
1111
10. E. coli có thể sinh trưởng trong môi trường chỉ chứa nguồn carbon là glucose. Như
vậy vi khuẩn đã tự tạo ra các hợp chất đường 5 carbon cho tổng hợp nucleotid, giải
thích?
Để sinh trưởng vi khuẩn cần năng lượng và vật chất để xây dựng nên tổ chức tế bào,
trong đó có DNA, RNA và các nucleotide cũng như các dẫn xuất của nó.
Năng lượng tạo thành từ glucose theo chuyển hóa glycolysis → TCA→ phosphoryl
hoa oxyhóa
Cảc hợp chất ribose 5-phosphate tiền chất để tổng hợp nucleotide được tổng hợp từ
Glc qua con đường pentose phosphate:
Giai đoạn 1: Glucose + ATP
glucose 6-phosphate
Giai đoạn 2: glucose 6-phosphate
ribose 5-phosphate
CHƯƠNG 6
Bài giải Chương 6: Chu trình Citric acid
1. Mặc dù oxy không trực tiếp tham gia vào chu trình citric acid nhưng chu trình này chỉ
bắt đầu khi có mặt oxy. Vì sao như vậy?
Trong phản ứng tạo acetyl-CoA từ pyruvate, NAD
+
được chuyển thành dạng NADH cung
cấp một ion hydride (:H
-
). Chính ion này sẽ là chất mang 2 electron theo chuỗi hô hấp và
chuyển đến phân tử oxy. Như vậy vai trò của oxy nhận điện tử và tái tạo lại NAD
+
cung cấp
cho chu trình citric acid để từ đó tổng hợp NADH chủ yếu trong chu trình TCA, do vậy hoạt
hóa chu trình này . Nếu như không có oxy, NADH tích lũy và chu trình TCA vì vậy dừng lại
1212
2. Tế bào eukaryote có thể sử dụng glucose (C
6
H
12
O
6
) và hexagonic acid (C
6
H
12
O
2
) như là
nguồn nhiên liệu cho hô hấp tế bào. Dựa trên cấu trúc phân tử, chất nào sẽ cho nhiều năng
lượng hơn khi đốt cháy (oxy hóa sinh học) chúng thành CO
2
và H
2
O?
Hexagonic acid là acid béo no (C
6
H
12
O
2
) : CH3-CH2-CH2-CH2- CH2- COOH
1 phân tử glucose chuyển hóa thành 32 ATP
1 phân tử hexagonic acid oxi hóa hóa beta (chương 7) thông qua 3 giai đoạn:
Giai đoạn-1: hoạt hóa acid béo tạo thành hexagonic-CoA
Hexagonic acid+ ATP + Co-SH → hexagonic-CoA + AMP +PPi
Như vậy tiêu tốn -1ATP
Giai đoạn 2: oxi hóa beta tạ, thành 3 phân tử Acetyl-CoA
2 lần oxi hóa beta tạo 3 acetyl-CoA thì tạo thành +2NADH và +2FADH2 tương đương
=[(n/2)-1]*(2,5+1,5)
= (6/2-1)x(2,5+1,5)= +8ATP
Giai đoạn 3: 1 phân tử Acetyl-CoA đi vào chu trình citric acid tạo thành 3NADH +
1FADH2 + ATP, tổng là 3*2,5 +1,5 +1 =10 ATP. Vậy, 3 phân tử acetyl-CoA tạo thành
11*3 = 30 ATP
Như vậy cả 3 giai đoạn là 37 ATP > 32 ATP của glucose
Lập công thức tính chung sẽ là
Tổng ATP của oxi hóa 1 acid béo no có n carbon số chẵn = (n/2-1)*4 + 5n -1
áp dụng cách tính này cho palmitic-CoA xem có đúng là 108 ATP không?
= (16/2-1)*4 + 5*16 =28 +80= 108
Nếu tính cả giai đoạn hoạt hóa palmitic thành palmitic-CoA thì =108-1=107 ATP
1313
Bằng cách này các em có thể tự lập công thức cho số ATP của acid béo có số carbon lẻ (nhớ
là 3 carbon cuối propionic-CoA thì tạo thành succinate-CoA) , hoặc cho dạng acid béo có
chứa nối đôi (phải tính thêm các phản ứng hydrogen hóa nối đôi có tiêu thụ/tạo ATP?)
Hoặc trong trường hợp hexagonic acid là acid béo no, do đó có thể ước lượng nhanh theo
ví dụ đã cho, palmitic acid, tính từ Palmitic-CoA thì acid này bị oxihóa beta tạo thành 8
Acetyl-CoA và tổng năng lượng là 108 ATP (chương 7) vậy hexgonic acid tạo thành 8
Acetyl-CoA năng lượng khoảng 108*3/8 = 40.5 ATP nếu trừ đi 1 ATP hoạt hóa CoA ban
đầu thì = 39.5 ATP gần đúng với 37 ATP
3. Những người bị bệnh phù beriberi do thiếu thiamine (Vitamin B1) sẽ có nồng độ pyruvate và
α-ketoglutarate cao trong máu đặc biệt sau bữa ăn giàu glucose. Những tác động trên có liên
hệ gì với sự thiếu thiamine?
Thiamine dưới dạng thiamine pyrophosphate là yếu tố thiết yếu tạo nên hoạt tính enzym
pyruvate dehydrogenase, enzym chuyển hóa pyruvate thành Acetyl –CoA tiền chất của chu
trình citric acid. Do đó, sự thiếu vitamine này làm giảm hoạt tính enzyme trên dẫn đến tích
lũy pyruvate.
Trong chu trình Kreb, α-ketoglutarate chuyển hóa thành succinyl-CoA nhờ enzym α-
ketoglutarate hydrogenas, mà enzym này có cấu hình và hoạt động tương tự như enzyme
pyruvate dehydrogenase do vậy thiếu hụt Thiamin còn gây giảm hoạt tính α-ketoglutarate
hydrogenase cho nên đồng thời gây tích lũy α-ketoglutarate.
4. Oxaloacetate được hình thành ở bước cuối cùng của chu trình citric acid bởi sự oxihóa L-
malate phụ thuộc NAD
+
. Không cần loại đi các chất biến dưỡng trung gian, sự tổng hợp
oxaloacetate từ acetyl-CoA chỉ dùng các enzyme và cofactor của chu trình citric acid có thể
xảy ra được hay không? Giải thích. Vì sao oxaloacetate bị mất mát trong chiều ngược lại của
chu trình?
1414
1515
Oxaloacetate có thể tổng hợp từ Acetyl-coA được với điều kiện phải giữ nguyên các
chất biến dưỡng trung gian và các enzym của từng phản ứng. đánh dấu 2 carbon của
Acetyl -CoA , qua chu trình 2 carbon này vẫn có mặt trong oxaloacetate.
Oxaloacetate bị mất đi theo chu trình ngược lại vì các phản ứng là thuận nghịch, nếu như nồng
độ 1 chất trung gian giảm giả sử malate hay sucinate thì oxaloacetate sẽ bị thiết hụt
CHƯƠNG 7 : LIPID
1. Điểm nóng chảy của những acid béo có 18 carbon như sau: stearic acid, 69.6
o
C; oleic
acid, 13.4
o
C; linoleic acid, -5
o
C; linolenic acid, -11
o
C.
a- Những đặc điểm nào về cấu trúc của những acid béo 18 carbon này liên quan
tới điểm nóng chảy của nó? Cung cấp một giải thích ở mức độ phân tử cho khuynh hướng
trên của điểm nóng chảy.
Điểm nóng chảy của acid béo phụ thuộc vào độ dài và mức độ không bão hòa của chuỗi
hydrocacbon.
Đối với các acid béo bão hòa hoàn toàn, các nguyên tử C liên kết đơn với nhau có thể
quay tự do, làm cho cấu trúc phân tử này mềm dẻo. Trạng thái bển nhât của các phân từ
này là trạng thái duỗi thẳng hoàn toàn, vì sự cản trở xảy ra giữa các phân tử kế cận
nhau là tối thiểu. Do đó, các phân tử này có thể tập hợp kế cận nhau chặt chẽ bởi các
liên kết van der Waals giữa 2 nguyên tử của 2 phân tử kế cận nhau. Điểm nóng chảy của
các phân tử này cao.
Đối với các acid béo không bão hòa, liên kết đôi của các nguyên tử C (cấu hình cis) bẻ
cong mạch hydrocarbon của phân tử acid béo. Các acid béo này sẽ không thể xếp chặt
chẽ cạnh nhau, liên kế giữa chúng yếu đi dẫn đến dễ phá vỡ các liên kết này. Điểm nóng
chảy của các phân tử này thấp.
b- Hãy vẽ tất cả các triacylglyceride được tạo thành từ glycerol, palmitic acid và
oleic acid. Sắp xếp chúng theo thứ tự tăng dần về điểm nóng chảy.
Glycerol có thể kết hợp với 3 phân tử palmitic acid (P) hoặc 3 phân tử oleic acid (O);
hoặc 3 phân tử của cả hai acid này tại nhóm –OH của nó để tạo thành các
tryacylglyceride. Các tryacylglyceride tạo được có thể là: OOO, PPP, OOP, OPO, POP,
OPP. Điểm nóng chảy phụ thuộc vào chiều dài và mức độ bão hòa của chuỗi
1616
hydrocacbon (càng bão hòa thì điểm nóng chảy càng cao), vì vậy thứ tự tăng dần về
nhiệt độ nóng chảy sẽ như sau:
OOO < OOP = OPO < OPP = POP < PPP (nhiệt độ nóng chảy cao nhất)
c- Các acid béo mạch nhánh được tìm thấy trong màng lipid của một vài vi khuẩn.
Sự có mặt của chúng làm tăng hay giảm tính lỏng của màng? Tại sao?
Các acid béo có mang mạch nhánh sẽ làm lỏng lẻo liên kết của các nhóm phân tử acid
béo vì chúng cản trở sự kết chặt các phân tử acid béo khác kế cận nó, dẫn đến làm tăng
tính lỏng lẻo của màng.
2. Một tiến trình phổ biến để làm sạch quần áo dính dầu mỡ là cho vào đó những sản
phẩm có chứa sodium hydroxide. Giải thích việc làm này
Dẩu mỡ (thành phần chính là các triacylglyceride đơn giản và phức tạp)và acid béo tác
dụng bởi NaOH trong phản ứng xà phòng hóa, tạo glycerol và các muối Natri của acid
béo (vd: sodium stearate) có khả năng tan trong nước vì vậy dẩu mỡ từ bị khử liên kết kị
nước gây bám dính trên vật liệu rắn và dễ dàng dễ bị rửa trôi.
3. Hãy sắp xếp theo thứ tự tăng dần độ hòa tan trong nước các glyceride từ palmitic acid
sau: triacylglycerol, diacylglycerol, monoacylglycerol
Triacylglycerol < Diacylglycerol < Monoacylglycerol
Vì các phân tử hydroxyl của glycerol và nhóm carboxyl của các acid béo liên kết ester
với nhau tạo các acylglycerol, do vậy số phân tử acid béo gắn vào glycerol sẽ quyết định
đến tính tan trong nước. Càng nhiều nhóm –OH cua glycerol bị liên kết thì các phân tử
acylglyceride này càng không phân cực, kỵ nước, vì thế khộng tan trong nước.
4. Trên cơ sở mỗi phân tử carbon, thành phần nào trên phân tử triacylglycerol sẽ cho
lượng năng lượng sinh học lớn nhất: phân tử glycerol hay acid béo? Hãy giải thích lựa
chọn của bạn dựa vào những hiểu biết về cấu trúc phân tử của triacylglycerol
Triacylglycerol khi thủy phân sẽ tạo thành Glyceryl và acid béo, mỗi thành phần này đều
có thể được chuyển hóa để thành năng lượng. Glycerol chuyển hóa thành 3-
phosphoglyceradehyde : -ATP + NADH + 3-P glyceraldehyde, chất này tiếp tuc chuyển
1717
hóa glycolysis và TCA rồi phosphoryl hóa oxy hóa thành 16 ATP như vậy tổng 1 glycerol
thành khoảng 18,5 ATP.
Còn 3 phân tử acid béo, thông thường có số carbon dài vì dụ như 16 carbon palmitic
acid đã cho 108 ATP, vậy 3 phân tử acid béo cho >300ATP
5. Trái với truyền thuyết, lạc đà không giữ nước trong các bướu của chúng, nơi mà thực
sự chúng tích lũy chất béo. Những chất béo dự trữ này đóng vai trò là nguồn cung cấp
nước cho lạc đà bằng cách nào?
Để đơn giản ta coi toàn bộ chất béo ở bướu lạc đà là tripalmitoylglycerol. Hãy tính thể
tích nước (bằng lit) lạc đà có thể tạo thành từ 1.0kg chất béo dự trữ.
Chất béo dự trữ trước sẽ bị thủy phân thành Glycerol + 3 acid béo
Quá trình oxy hóa một acid béo sẽ có 3 giai đoạn
1/ hoạt hóa CoA→acidbéo-CoA
2/ oxy hóa beta → nhiều acetyl-CoA
3/ các acetyl-CoA đi vào chu trình citric tạo thành CO2 và H2O và các phân tử ATP
Tripalmitoylglycerol bị thủy giải thành các palmitoyl-CoA và glycerol. Các acid béo sẽ
bị oxy hóa theo phương trình tổng quát sau:
Palmitoyl-CoA + 23O
2
+ 108P
i
+ 108ADP → CoA + 108ATP + 16CO
2
+ 23H
2
O
Từ đó, với tripalmitoylglycerol (MW: 806 g/mol) khối lượng 1kg thì có số mol của
palmitoyl-CoA là: 1000/806*3= 3,722 mol.
Theo phương trình trên số mol nước sẽ là: 23* 3,722= 85,6 mol.
Lượng nước tạo thành từ 1kg chất béo dự trữ: =85,6* 18= 1541 g ≈ 1.5 l
6. Một vài vi sinh vật thuộc giống Nocardia và Pseudomonas có thể sống được trên môi
trường chỉ có hydrocarbon làm nguồn dinh dưỡng chính. Những vi khuẩn này oxy hóa
các hydrocarbon béo mạch thẳng như octane thành các carboxylic acid tương ứng:
CH
3
(CH
2
)
6
CH
3
+ NAD
+
+ O
2
→ CH
3
(CH
2
)
6
COOH + NADH + H
+
Những vi khuẩn trên có thể được sử dụng để xử lý các vụ tràn dầu như thế nào? Điều gì
sẽ là nhân tố giới hạn cho hiệu quả của tiến trình này?
Những vi khuẩn trên có thể dung để xử lý các vụ tràn dầu một cách trực tiếp vì chúng có
thể sử dụng các hydrocarbon béo mạch thẳng chuyển hóa thành acetyl-CoA vào chu
trình TCA để tạo năng lượng chính. Từ đây Glc có thể được tạo thành theo con đường
gluconeogenesis, và Ribose-5-phosphate được tạo thành từ con đường pentophosphate
1818
Tuy nhiên, một số giới hạn khi xử lý theo hướng này hạn chế về sự tiếp xúc giữa enzyme
và cơ chất (enzyme ưa nước, cơ chất hydrocarbon béo mạch thẳng kỵ nước);cần quan
tâm đến các nguồn dinh dưỡng khác như nitrogen, phosphate. . và điều kiện hô hấp là
oxy.
7. Sử dụng các kiến thức đã biết về sự sinh tổng hợp acid béo, hãy giải thích các kết quả
thí nghiệm quan sát được sau:
a- Bổ sung [
14
C]-acetyl-CoA được đánh dấu phóng xạ toàn bộ vào một phân đoạn
hòa tan của gan thì palmitate sinh ra đều được đánh dấu phóng xạ toàn bộ.
b- Tuy nhiên, nếu chỉ bổ sung [
14
C]-acetyl-CoA được đánh dấu phóng xạ toàn bộ
dưới dạng vết trong phân đoạn hòa tan của gan với lượng thừa malonyl-CoA không đánh
dấu thì palmitate sinh ra chỉ được đánh dấu ở C-15 và C-16.
a- Trong môi trường chỉ bổ sung acetyl-CoA thì các malonyl-CoA sinh ra đều có
nguồn gốc từ acetyl-CoA. Do đó, khi bổ sung toàn bộ [
14
C]-acetyl-CoA vào phân
đoạn hòa tan của mô gan thì sẽ sinh ra các [
14
C]-malonyl-CoA và các acid béo
tạo thành cũng được đánh dấu phóng xạ toàn bộ.
b- Trong khi nếu chỉ bổ sung [
14
C]-acetyl-CoA dưới dạng vết và các malonyl-CoA có
sẵn trong môi trường thì chỉ có carbon C-15 và C-16 của palmitate là được đánh
dấu vì hai vị trí này có nguồn gốc từ hai carbon methyl và carboxyl của acetyl-
CoA.
8. Hãy viết chuỗi phản ứng sinh tổng hợp palmitate ở gan chuột bắt đầu từ acetyl-CoA ty
thể và NADPH, ATP, CO
2
từ cytosol.
Trong phản ứng tổng hợp acid béo,
1- acetyl-CoA được carboxyl hóa (gắn thêm CO2) → malonyl-CoA.
7 acetyl-CoA + 7CO2 + 7ATP → 7 malonyl-CoA +7ADP + 7Pi
2 Tổng hợp acid béo: từ malonyl-CoA + acetyl-CoA thành palmityl-CoA trong đó có sử
dụng NADPH + H
+
.
Acetyl-CoA + 7malonyl-CoA +14 NADPH + 14H
+
→ Pamitate + 7CO2 + 8CoA +
14NADP
+
+ 6H
2
O
Phản ứng tổng quát
1919
8Acetyl-CoA + 14NADPH + 14H
+
+ 7ATP → palmitate + 14NADP
+
+ 7ADP + 7P
i
+
8CoA + 6H
2
O
2020